
Neurosciences/La vision
Pour commencer ce chapitre, nous allons faire un petit aparté sur l’œil. Une bonne partie de l’œil n'appartient pas au système nerveux, mais il est bon de faire un petit rappel sur son anatomie, histoire de se remettre dans le contexte. L’œil est entouré par un tissu de couleur blanche, la sclérotique, qui compose le blanc des yeux. Entourée par cette sclérotique, on trouve une sphère remplie d'une substance, l'humeur vitrée. La lumière entre dans l’œil à travers la cornée et traverse un liquide, l'humeur aqueuse. Elle arrive alors sur le cristallin, où la lumière est concentrée sur le fond de l’œil : le cristallin sert à faire converger les rayons lumineux qui proviennent d'un même endroit en un même point du fond de l’œil. En somme, ce cristallin est l'équivalent de la lentille focale d'un appareil photographique, mais pour l’œil. Il peut se déformer sous l'action des muscles ciliaires : cela permet d'adapter la convergence des rayons lumineux en fonction de la distance de l’objet regardé. Au-dessus du cristallin, on trouve une pupille, un tissu qui peut se contracter ou se dilater pour contrôler la quantité de lumière qui entre dans l’œil. Cette dilatation ou contraction est effectuée par des muscles dédiés. La lumière est alors captée et transformée en potentiels d'action au niveau de la rétine, au fond de l’œil.

Le système visuel : aperçu
[modifier | modifier le wikicode]Le système visuel est assez complexe, aussi allons-nous commencer par une vue d'ensemble. Dans les grandes lignes, les informations visuelles sont captées par la rétine, avant d'être transmises par le nerf optique. Les deux nerfs optiques se croisent au niveau du chiasma optique, avant de poursuivre leur chemin. Les nerfs optiques font synapse sur le thalamus, au niveau des corps genouillés latéraux. Puis, le thalamus relaie les informations visuelles au cortex occipital, dédié à la vision, où les informations visuelles subissent des traitements visuels élémentaires (détection de la forme, des couleurs, ...). Les aires suivantes, à savoir le cortex temporal et pariétal, s'occupent d'identifier les objets perçus ou leur mouvement : elles donnent un sens à ce qui est perçu. Pour résumer, les informations sont captées par la rétine, envoyées au thalamus par un ensemble de nerfs (nerfs optiques, chiasma optique et tractus optiques), puis transmises au cortex occipital par les radiations optiques.

Les voies parvocellulaire et magnocellulaire
[modifier | modifier le wikicode]Évidemment, rétine, thalamus et cortex visuel ont chacun une contribution différente au traitement de la vision. Le gros du travail se fait dans le cortex visuel, mais aussi dans la rétine, le corps genouillé latéral du thalamus ne semblant pas faire beaucoup de traitements visuels complexes. Voir quelque chose demande de traiter un grand nombre d'informations différente : la couleur, la luminosité, le contraste, etc. À partir de ces informations basiques, le cerveau peut alors détecter les formes et contours d'un objet, détecter un mouvement quelconque, reconnaitre un objet, traiter ses couleurs, etc. Pour simplifier, on peut décomposer les stimulus visuels en trois composantes : leur couleur, leur forme et leur mouvement. Cette distinction est très importante, car le traitement des couleurs est séparé du reste sur une grande partie du système visuel. La transmission des couleurs est séparée de la transmission des autres stimulus, dans le sens où les axones sont ségrégés, les aires cérébrales impliquées sont différentes, etc. Les cellules de la rétine en charge de la perception des couleurs sont différentes des cellules en charge du reste, par exemple. De même, dans le corps genouillé latéral, les couches neuronales en charge des couleurs sont différentes de celles en charge du mouvement et des formes. Et c'est la même chose dans le cortex visuel, jusqu’à un certain point.
Pour faire court, il existe trois voies de transmission/traitement :
- une voie magnocellulaire, qui transmet les informations sur leur mouvement, comme leur vitesse ;
- une voie parvocellulaire, qui transmet les informations sur la couleur des objets ;
- une voie koniocellulaire, très mal connue à l’heure où j'écris ces lignes.
La voie parvocellulaire est de loin celle qui contient le plus d'axones et de neurones. Elle regroupe pas moins de 80% des fibres nerveuses et axones dédiés à la vision, les autres voies se contentant des 20% restants. Elle transmet les informations sur les couleurs, mais aussi sur la vision noir et blanc quand le contraste est élevé. La voie magnocellulaire, quant à elle, prend en charge la détection des mouvements, mais aussi la vision noir et blanc quand le contraste est faible. La différence tient aussi dans la taille des fibres : les fibres de la voir magnocellulaire sont bien plus grosses que celle de la voie parvocellulaire. En conséquence, la voie magnocellulaire a une vitesse de transmission assez rapide, et elle permet de réagir rapidement aux stimulus visuels (ce qui colle bien avec la perception des mouvements). À l'inverse, la voie parvocellulaire est plus lente, avec une vitesse de transmission nerveuse bien moins rapide, ce qui n'est pas si désavantageux pour la perception des couleurs.
Pour résumer, la voie parvocellulaire est très sensible aux couleurs, mais peu à la luminosité, au contraste, au mouvement et aux formes. Elle est donc naturellement en charge de la perception des couleurs. Elle a une bonne acuité visuelle, mais est lente, met du temps à réagir à un stimulus. Par contre, la voie magnocellulaire est bien plus rapide, mais a une acuité nettement plus faible. Elle est sensible au contraste et à la luminosité, mais très peu aux couleurs. Ces caractéristiques font qu'elle est en charge de la détection des formes et du mouvement.
 |
 |
Les champs récepteurs
[modifier | modifier le wikicode]Si on prend un neurone, peu importe sa position dans les voies visuelles, il va réagir quand on soumet la rétine à un stimulus. Tous ne vont pas réagir aux même stimulus : certains vont réagir à la présence de lumière, d'autre à un stimulus mobile, d'autres à une couleur en particulier, etc. Mais surtout, ils vont réagir pour une portion assez précise de la rétine. Par exemple, prenons un neurone du cortex visuel primaire : il ne va réagir qu'aux stimulus présentés sur une petite portion de la rétine, mais ne va pas réagir aux autres. Comme autre exemple, un neurone du thalamus pris au hasard va réagir quand on illumine une petite portion de la rétine, mais rester inerte quand cette même portion est laissée dans l'obscurité. Dans tous les cas, cette portion de la rétine est appelée le champ récepteur du neurone considéré. Notons que la portion dépend du neurone considéré et que les champs récepteurs de deux neurones sont différents.

La taille d'un champ récepteur varie selon la position dans les voies visuelles et la taille augmente au fur et à mesure qu'on se rapproche du cerveau. Les neurones de la rétine ont des champs récepteurs très petits, ceux du thalamus un champ récepteur moyen et ceux du cortex un champ récepteur bien plus gros. Il en est de même à l'intérieur du cerveau : les champs récepteurs sont plus petits dans les aires visuelles dites primaires (les portes d'entrée de la vision dans le cerveau), mais plus gros dans les aires secondaires où s'effectue le traitement de l'information. Le schéma ci-contre montre que la taille des champs récepteurs augmente quand on passe d'une aire visuelle à la suivante, à l'intérieur du cerveau.
Tout cela trahit le fait que les informations visuelles sont intégrées et deviennent de plus en plus abstraites au fur et à mesure que l'on progresse dans les voies visuelles. Si les neurones de la rétine traitent des signaux locaux et ne font que des traitements de bas-niveau (amplification du contraste, détection des couleurs d'un "pixel" de la rétine), les aires finales traitent des informations plus conceptuelles (reconnaissance des objets, ...). Qui plus est, on peut remarquer que les neurones deviennent de plus en plus sélectifs quand on progresse dans les voies visuelles. Si les neurones proches de la rétine réagissent à des stimulus simples (couleur, mouvement, forme), ceux des aires de haut-niveau réagissent à des stimulus de plus en plus précis. Dans la rétine, le stimulus classique classique qui fait réagir les neurones est un disque de lumière de la même taille que le champ récepteur. Cela ne change pas beaucoup dans le corps genouillé latéral, qui sert de relai et n'effectue que peu de traitements. Dans le cortex visuel primaire, certains neurones vont réagir uniquement à des colonnes orientées dans une certaine direction. Et dans le cortex visuel secondaire, les neurones ne réagissent qu'à des stimulus complexes, comme des visages ou des objets bien précis. Par exemple, certains neurones ne s'activent que quand on leur présente une photographie d'un proche, une image de son visage, ou quelques stimulus ressemblants.
La rétine et les photorécepteurs
[modifier | modifier le wikicode]La rétine est une couche de neurones qui tapisse le fond de l’œil. Elle est alimentée en nutriments et protégée par une couche choroïde. Celle-ci est en continuité avec le nerf optique, ce dernier étant composé des axones des neurones de la rétine. Il faut cependant noter que les neurones de la rétine ne sont pas considérés comme appartenant au système nerveux périphérique. Les raisons sont multiples. La première est que lors du développement du système nerveux, la rétine se forme par protrusion du système nerveux central (plus précisément, il s'agit d'une excroissance du diencéphale). De plus, les cellules gliales de la rétine sont identiques à celles qu'on trouve dans le système nerveux central, et non celles présentes dans le système nerveux périphérique (relisez le chapitre sur les cellules gliales et les neurones si vous avez oublié les différences).
La rétine est composée de plusieurs couches de neurones, ce qui en fait un organe relativement complexe. Elle effectue de nombreux traitements sur la lumière qui lui parvient, comme ceux liés au contraste. Dans les grandes lignes, on trouve plusieurs types de neurones dans la rétine :
- une couche de photorécepteurs qui captent la lumière et émettent des potentiels d'action en fonction de la luminosité ou de la couleur perçue ;
- une couche de neurones bipolaires ;
- une couche de neurones ganglionnaires ;
- on trouve aussi des neurones annexes dans chaque couche : des cellules amacrines et horizontales.
Les photorécepteurs
[modifier | modifier le wikicode]Les photorécepteurs, aussi appelés cellules photoréceptrices, sont des récepteurs sensoriels capables de capter la lumière dans un certain intervalle de luminosité ou de couleur et la transforment en potentiels d'action. Pour cela, ces cellules contiennent des pigments, des molécules qui réagissent à la lumière, qui portent eux aussi le nom de photorécepteurs par abus de langage. Lorsque ces photorécepteurs reçoivent de la lumière, elles déclenchent une cascade de réactions chimiques qui modifient le potentiel à la surface du neurone, pouvant déclencher un potentiel d'action. Toutes les cellules photoréceptrices sont composées de deux segments :
- un segment interne avec le corps cellulaire et les mitochondries ;
- un segment externe avec les photorécepteurs.
Les deux types de photorécepteurs : cônes et bâtonnets
[modifier | modifier le wikicode]Mais malgré leurs ressemblances, les photorécepteurs peuvent être classés en deux catégories :
- les cônes perçoivent la lumière intense, permettant de voir les couleurs ;
- les bâtonnets perçoivent les faibles luminosités et permettent la vision de nuit.
Cônes et bâtonnets en sont pas sensibles au même intervalle de luminosité : les cônes captent bien les luminosités importantes mais ce n'est pas le cas des bâtonnets. Quand la lumière est peu intense, seuls les bâtonnets sont actifs : on parle de vision scotopique. C'est cette vision qui permet de voir dans une nuit profonde. Et d'ailleurs, vu que les bâtonnets ne captent pas les différences de couleurs, la vision nocturne est essentiellement en gris, noir et blanc : après tout, le proverbe dit que la nuit, tous les chats sont gris. Vient ensuite un intervalle de lumière où cônes et bâtonnets sont actifs : c'est la vision mésoscopique, qui permet de voir lors d'un clair de lune ou d'un crépuscule. Vient ensuite la luminosité intense, typique d'une journée normale : seuls les cônes sont actifs, donnant une vision photopique.


Cônes et bâtonnets sont tous deux composés de trois parties : un segment externe et un segment interne, suivis par une terminaison synaptique. Le segment interne est le lieu où les pigments photorécepteurs sont régénérés, par toute une série de réactions chimiques. Le segment externe est le lieu où se trouvent les molécules photoréceptrices, les pigments, et c'est dans ce segment que la lumière est transformée en potentiels d'action. Le segment externe a une forme cylindrique pour les bâtonnets, mais une forme de cône pour les cônes (d'où leurs noms). Chaque bâtonnet contient une pile de disques empilés les uns sur les autres, comme des comprimés dans un tube de médicaments. C'est sur la membrane externe des disques que se trouvent les molécules du pigment photorécepteur, la rhodopsine.
Les bâtonnets contiennent un pigment appelé rhodopsine, que nous avons vu dans les premiers chapitres de ce cours. On a vu que celle-ci est uniquement sensible à la luminosité générale, mais pas aux couleurs. En conséquence, les bâtonnets ne captent pas les couleurs, les pigments visuels qu'ils contiennent étant sensibles à l'ensemble du spectre de la lumière visible.
Ce n'est pas le cas des cônes, dont le segment externe contient un pigment visuel spécialisé dans la perception des couleurs : l'iodopsine. Elle existe en trois versions, qui réagissent à des couleurs différentes : une pour le vert, une autre pour le rouge et une autre pour le bleu. Ainsi, chaque couleur perçue est un mélange de ces trois couleurs dans des proportions différentes. Si jamais certains types de cônes ne sont pas présents (les cônes verts, par exemple), le sujet est daltonien : il ne peut pas percevoir une des trois couleurs primaires. Il faut bien signaler que s'il existe des sujets daltoniens qui ne voient pas le rouge, il existe aussi des sujets qui ne voient pas le vert ou le bleu sans difficultés avec les autres couleurs primaires. Généralement, l'origine de ce trouble est génétique.
Notons cependant que la génération du potentiel d'action a lieu sur la membrane du segment externe, pas dans les disques ! Il y a donc un intermédiaire qui se transmet des disques vers la membrane. Cette intermédiaire n'est autre que de la Guanosine MonoPhosphate Cyclique (GMPc). Nous en avions déjà parlé dans le chapitre sur les récepteurs sensoriels, aussi nous ne ferons que quelques rappels dans ce qui suit. La GMPc agit sur des canaux ioniques au sodium, en les ouvrant. Dans l'obscurité, la concentration en GMPc est maximale et le neurone a ses canaux sodium ouverts : le sodium rentre dans le neurone comme dans un moulin. L'entrée d'ions sodium est compensée par l'expulsion d'ions potassium, jusqu’à atteindre un point d'équilibre électrochimique. Une fois exposé à la lumière la production de GMPc est fortement réduite. La teneur en GMPc n'est plus suffisante pour garder les canaux sodium ouverts, ce qui fait qu'ils se ferment. Les ions sodium étant chargés positivement, cela fait moins d'entrée de charges positives dans le bâtonnet et donc une augmentation des charges négatives. Le résultat est que le photorécepteur se polarise donc encore plus qu'il ne l'était déjà : sa tension devient encore plus négative qu'à l'obscurité. Cette hyper-polarisation se transmet jusqu’à la terminaison synaptique et se transmet jusqu’aux couches de la rétine. Ce processus de transduction est illustré ci-dessous.
Fovéa et tache aveugle
[modifier | modifier le wikicode]
Les cônes et bâtonnets ne sont pas répartis uniformément sur la rétine. En effet, les cônes sont surtout rassemblés à un endroit bien précis de la rétine : la macula lutea. Au centre de celle-ci, la concentration en cônes augmente progressivement au fur et à mesure qu'on se rapproche du centre, où il existe une zone dépourvue de bâtonnets, la fovéa. Celle-ci permet donc la vision centrale, c'est là que l'on amène les images des objets proches de la direction du regard (du point de fixation).
En comparaison, les bâtonnets sont dispersés sur toute la rétine. Ils s'occupent donc de la vision périphérique. Il y a juste une zone de la rétine qui est sans photorécepteurs et ne peut pas percevoir de lumière. C'est l'endroit de la rétine qui surmonte le nerf optique. On l'appelle la tache aveugle.

Les couches et cellules de la rétine
[modifier | modifier le wikicode]En dessous de la couche de photorécepteurs, on trouve deux couches, respectivement formées de cellules bipolaires et de cellules ganglionnaires. Sur les synapses entre les trois couches de neurones, on trouve des cellules amacrines et horizontales à la fonction encore assez obscure. Ainsi, on trouve plusieurs couches dans la rétine :
- celle des photorécepteurs (elle-même coupée en deux, avec la présence de segments externes et internes) ;
- celle des synapses entre photorécepteurs et cellules bipolaires, qui contient des cellules horizontales ;
- celle des cellules bipolaires ;
- celle des synapses entre cellules ganglionnaires et cellules bipolaires, qui contient des cellules amacrines ;
- celle des cellules ganglionnaires ;
- celle des axones des cellules ganglionnaires, qui convergent vers le nerf optique.

Si on omet les photorécepteurs, les cellules ganglionnaires et bipolaires sont les plus étudiées par les scientifiques, les cellules amacrines et horizontales étant moins bien connues. Il faut dire que la rétine contient bien plus de cellules bipolaires et ganglionnaires que de cellules horizontales et amacrines.
Les cellules bipolaires
[modifier | modifier le wikicode]Pour rappel, un neurone bipolaire est un neurone avec une seule dendrite et un seul axone. Dans la rétine, ils servent de relais entre les photorécepteurs et une couche de cellules ganglionnaires. Ces dernières reçoivent des informations en provenance d'un ou de plusieurs neurones bipolaires. Il faut noter que les bâtonnets et les cônes ne sont pas reliés de la même manière aux neurones bipolaires. Un neurone bipolaire de la fovéa n'est relié qu'à un seul cône, alors qu'ils sont toujours reliés à plusieurs bâtonnets dans le reste de la rétine. La conséquence est que la vision centrale (captée par la fovéa) est plus précise, plus fine, que la vision périphérique.
Les cellules ganglionnaires
[modifier | modifier le wikicode]Il existe plusieurs types principaux de cellules ganglionnaires et nous allons surtout nous intéresser aux petites cellules ganglionnaires (midget cells) et les grandes cellules ganglionnaires (cellules parasol). Comme leur nom l'indique, elles n'ont pas la même taille, mais ce n'est pas ce qui est le plus important. Si on peut classer les cellules ganglionnaires en fonction de leur taille, cette classification n'est pas très pertinente. À la place, les scientifiques préfèrent classer les cellules ganglionnaires selon leurs projections (les aires cérébrales vers lesquelles elles envoient leur axones) et selon leur fonction. Cela permet de distinguer plusieurs types de cellules ganglionnaires :
- Les midget cells, aussi appelés P cells, ou encore petites cellules ganglionnaires ;
- Les parasol cells, aussi appelés M cells, ou encore grandes cellules ganglionnaires ;
- Les cellules bistratifées, aussi appelés K cells, ou encore cellules koniocellulaires ;
- Les cellules ganglionnaires photosensibles, impliquées dans la régulation veille-sommeil ;
- et quelques autres types de cellules moins bien connus.
Les trois premiers types sont un peu à part des cellules ganglionnaires photosensibles. Il faut dire que les cellules P, M et K sont impliquées dans la vision et ont des projections similaires. Elles émettent majoritairement leurs axones en direction du thalamus, et plus précisément des corps genouillés latéraux. Notons que les cellules parasol et les midget cells envoient leurs axones dans des zones séparées du thalamus et du cortex visuel. Elles correspondent à la voie parvocellulaire pour les cellules P et la voie magnocellulaire pour les cellules M, ainsi qu'à la voie koniocellulaire pour les cellules K (d'où leur nom : P pour parvocellulaire, M pour magnocellulaire et K pour koniocellulaire). Pour résumer, les petites cellules ganglionnaires se chargent de la perception des couleurs, alors que les grandes s'occupent de la perception de la luminosité, des formes, du contraste, du mouvement et de quelques autres stimulus visuels.

Les midget cells et parasol cells se connectent différemment aux cônes de la rétine. Les midget cells se connectent à des cônes de couleur très différentes et font une sommation de leurs émissions. Elles ne jouent pas de rôle dans la perception des couleurs, vu qu'elles intègrent les stimulus provenant de cônes de couleurs très différentes, leur rôle étant surtout la perception du mouvement ou des formes. Les cellules parasol ne font synapse qu'avec un petit nombre de cônes, tous spécialisés dans une ou deux couleurs. Elles sont donc assez sélectives et ne répondent qu'à une couleur bien précise. Par exemple, telle cellule parasol répond à un mélange de couleur rouge et verte (du jaune, donc), telle autre seulement à la couleur rouge accompagnée ou non de vert et/ou de bleu, telle autre à la présence de couleur rouge en l'absence de couleur verte, etc.

Les cellules ganglionnaires photosensibles, aussi appelées intrinsically photosensitive retinal ganglion cells ('ipRGCs'), sont un peu particulières. Elles n'ont aucun rôle dans la perception visuelle consciente proprement dite, mais sont impliquées dans la régulation de l'horloge biologique. Elles contiennent un pigment photosensible, la mélanopsine, qui leur permet de capter la lumière. Elles servent de photorécepteurs par elles-mêmes et n'ont pas besoin d’afférences des cônes ou bâtonnets (même si elles ont quelques synapses mineures avec). Elles réagissent à la lumière en se dépolarisant, ce qui est l'exact inverse des autres photorécepteurs : elles émettent des potentiels d'action quand elles sont stimulées par la lumière, alors que les autres photorécepteurs cessent d'en émettre.
Les cellules ganglionnaires photosensibles n'émettent pas le moindre axone vers le thalamus ou le cortex, mais font synapse dans l'hypothalamus, au niveau du noyau supra-chiasmatique. Cette aire cérébrale est en charge de la synchronisation de notre horloge biologique : elle contrôle les cycles hormonaux, les cycles veille-sommeil, ainsi que d'autres fonctions biologiques qui doivent se synchroniser sur le cycle jour-nuit ou les saisons. Les cellules ganglionnaires captent le niveau de luminosité, ce qui permet au noyau supra-chiasmatique de savoir s'il fait jour ou nuit. Elles jouent aussi un rôle dans la commande de certains réflexes, comme la dilation/contraction des pupilles, indépendamment de leur rôle dans l'horloge biologique.
Les cellules amacrines
[modifier | modifier le wikicode]Les cellules amacrines se situent dans la couche entre les cellules bipolaires et les cellules ganglionnaires. Elles forment des synapses dendrite-dendrite entre cellules ganglionnaires, qui sont de nature inhibitrices. Elles émettent majoritairement des neurotransmetteurs inhibiteurs, avec quelques neurotransmetteurs excitateurs de manière plus limitée. Classifier les cellules amacrines a été un vrai challenge, et il n'est pas certain que les classifications actuelles soient exemptes de défauts. Il faut dire que les cellules amacrines ont une diversité en terme de forme qui est assez importante. Les premières classifications morphologiques dépassaient la dizaine de sous-types de cellules amacrines, et certaines identifiaient près d'une centaine de types. Mais il était difficile de savoir si cette diversité morphologique reflète une réelle différence entre des types différents ou s'il s'agit de variations non-pertinentes d'un même sous-type de cellule.
.png)
De nos jours, les classifications des cellules amacrines se basent surtout sur les neurotransmetteurs qu'elles émettent/captent. Par exemple, on sait que certaines synthétisent du GABA, d'autres la glycine, que d'autres sont dopaminergiques, d'autres synthétisent de l'acétylcholine, etc. Dans les grandes lignes, on distingue trois types principaux de cellules amacrines : celles qui sont GABAergiques, celles qui sont glycinergiques et celles qui ne sont ni l'un ni l'autre, chaque type ayant de nombreux sous-types. Les cellules GABAergiques sont de loin les plus nombreuses, suivie par les cellules glycinergiques, et enfin par les cellules neutres (près de 15% des cellules amacrines ne sont ni GABAergiques, ni glycinergiques). Ces différences de neurotransmetteurs semblent correspondre à des différences entre terme de forme, notamment au niveau de l'arborescence dendritique. Mieux : ces différences semblent refléter des différences fonctionnelles, toutes les cellules amacrines ne servant pas à la même chose.
Les champs récepteurs de la rétine
[modifier | modifier le wikicode]Diverses expériences ont montré que les cellules ganglionnaires captent de la lumière sur une portion relativement limitée de la rétine, appelée le champ récepteur de la cellule ganglionnaire. Pour faire simple, le champ récepteur d'une cellule ganglionnaire est composé de l'ensemble des photorécepteurs qui innervent une cellule ganglionnaire. Pour rappel, le concept de champ récepteur ne vaut pas que pour les cellules ganglionnaires et on peut l'étendre pour toute cellule des voies visuelles.
La taille d'un champ récepteur ganglionnaire dépend de la connectivité entre photorécepteurs, cellules bipolaires et cellules ganglionnaires. Dans la fovéa, la connectivité est très faible : un cône fait synapse avec une seule cellule bipolaire, qui lui-même fait synapse avec une seule cellule ganglionnaire. Le champ récepteur est alors limité à un seul photorécepteur et est donc très petit. En-dehors de la fovéa, les choses changent. Chaque bâtonnet fait synapse avec plusieurs cellules bipolaires, qui eux-même font synapse avec beaucoup de cellules ganglionnaires. Le champ récepteur d'un bâtonnet est donc bien plus grand et contient environ 125 photorécepteurs. On devine que la vision liée aux bâtonnets est moins fine que celle liée aux cônes. C'est pour cela que la vision de nuit est plus floue que la vision de jour : c'est lié à cette différence d'acuité visuelle entre bâtonnets et cônes.
Précisons cependant que la finesse de la vision est liée non seulement à la taille des champs récepteurs ganglionnaires, mais aussi à la manière dont ils sont répartis. Pour les bâtonnets, les champs récepteurs ganglionnaires sont si gros qu'ils se recouvrent : il n'y a pas de portion de la rétine qui ne fasse pas partie de plusieurs champs récepteurs ganglionnaires. Par contre, les champs récepteurs ganglionnaires des cônes ne se recouvrent pas du tout, tant ils sont petits.
Les cellules ganglionnaires ne réagissent pas aux même stimulus. Certaines cellules ganglionnaires réagissent au mouvement, d'autres à la présence de lumière, d'autres à certains stimulus, etc. Dans les grandes lignes, on peut distinguer approximativement deux grandes catégories : les cellules qui réagissent aux mouvements et celles qui réagissent à une couleur ou un mélange de couleurs. Tout cela devrait vous rappeler la différence entre voie magnocellulaire et parvocellulaire. Dans ce qui suit, nous allons parler des cellules qui amplifient le contraste, puis de celles qui captent les mouvements.
Les cellules d'amplification du contraste
[modifier | modifier le wikicode]
Le champ récepteur d'une cellule ganglionnaire a une forme grossièrement circulaire. Il est composé de deux portions : un centre en forme de disque et une périphérie en forme d'anneau juste autour. Et la cellule réagit différemment selon que l'on illumine son centre ou sa périphérie. Pour faire simple, elles réagissent aux différences de contraste entre le centre et la périphérie du champ récepteur. Une illumination complète du champ récepteur entraine souvent une réponse assez faible, avec une émission légère de potentiels d'action. Par contre, laisser le champ récepteur dans l'obscurité fait que la cellule reste inactive. Il faut que le centre et la périphérie captent une intensité différente pour que la cellule ganglionnaire réagisse et émettent une bouffée de potentiels d'action. Pour résumer, ces cellules ganglionnaires sont capables d'amplifier le contraste, à savoir la différence de luminosité entre deux points rapprochés. Cela fait croire au cerveau que le contraste est plus ou moins important que ce qui est réellement perçu.
Pour rentrer plus dans le détail, il faut faire la différence entre cellules ganglionnaires à centre ON et à centre OFF. Les cellules à centre ON sont stimulées par leur centre, mai inhibées par leur périphérie. Elles émettent fréquemment des potentiels d'action quand le centre du champ récepteur est éclairé, mais que sa périphérie ne l'est pas. Les cellules à centre OFF font l'inverse : il faut que le centre soit nettement moins éclairé que la périphérie pour qu'elles émettent des potentiels d'action. Les deux types de cellules sont en nombre presque égal, ce qui fait que chaque portion de la rétine contient les deux types de récepteurs. Pour résumer, si le centre est plus éclairé que la périphérie ou au contraire moins éclairé, elles émettent des potentiels d'action. Les cellules à centre ON et OFF réagissent donc au contraste entre le centre et la périphérie.

Ce comportement vient de l'interaction entre cellules ganglionnaires, bipolaires et horizontales. On classe les synapses en deux types : la synapse entre cellule bipolaire et cellule ganglionnaire est appelée la voie directe, alors que les synapses qui font intervenir une cellule horizontale forment la voie indirecte. Les cellules bipolaires sont en contact direct avec les photorécepteurs situées au centre du champ récepteur, et en contact indirect avec ceux de sa périphérie par l'intermédiaire des cellules horizontales. Pour le dire autrement, le centre innerve la cellule ganglionnaire par la voie directe, alors que la périphérie passe par la voie indirecte. On devine donc que les cellules ganglionnaires à centre ON forment des synapses excitatrices avec les cellules bipolaires, mais aussi des synapses inhibitrices avec les cellules horizontales. Et inversement, pour les cellules à centre OFF, les synapses sont inhibitrices avec les cellules bipolaires et excitatrices avec les cellules horizontales.

Les cellules de détection du mouvement
[modifier | modifier le wikicode]À côté des cellules ganglionnaires précédentes, on peut aussi mentionner l'existence d'un autre type de cellule ganglionnaires, qui réagissent au mouvement. Plus précisément, elles réagissent aux mouvements qui vont dans une direction bien précise, appelée direction préférentielle. Pour simplifier, un stimulus se déplaçant dans la direction préférentielle active fortement la cellule ganglionnaire, alors qu'elle ne réagit pas à un stimulus allant dans la direction opposée, appelée direction nulle. Pour aller plus dans le détail, leur comportement est similaire à celui des cellules ganglionnaires précédentes. Si on illumine totalement leur champ récepteur, elles émettent des potentiels d'action de manière erratique et à une fréquence assez basse. Mais quand le stimulus lumineux se déplace dans la direction préférentielle, elles émettent une bouffée continue de potentiel d'action à haute fréquence. Inversement, un stimulus qui se déplace dans la direction inverse de la direction préférentielle (dans la direction nulle, stoppe totalement l'activité de la cellule ganglionnaire.
L'origine de ce comportement est encore mal comprise, mais elle est relativement bien décrite par le modèle de Harlow et Levick, que nous allons expliquer rapidement dans ce qui suit. Pour simplifier, certains photorécepteurs vont exciter la cellule ganglionnaire, alors que d'autres vont l'inhiber, comme pour les cellules ganglionnaires d'amplification du contraste. Sauf qu'ici, les photorécepteurs excitateurs et inhibiteurs sont répartis d'une manière bien précise, avec tous les photorécepteurs excitateurs d'un côté et les inhibiteurs de l'autre. Cette anisotropie du champ récepteur se traduit par l'existence d'une direction privilégiée. Lorsqu'un stimulus se déplace dans cette direction privilégiée, il va d'abord activer les photorécepteurs excitateurs, avant d'activer les inhibiteurs. Et lorsqu'il se déplace dans la direction nulle, ce sera l'inverse. Une illumination globale activera tous les photorécepteurs en même temps, ce qui donnera une réaction intermédiaire. Cela permet de rendre compte du comportement de la cellule ganglionnaire, sous une condition : les signaux inhibiteurs sont plus lents que les excitateurs et se propagent plus lentement dans les dendrites et axones. Ce faisant, pour un stimulus allant dans la direction nulle, les signaux inhibiteurs partis en premier pourront rattraper les signaux excitateurs et les annuler. Et pour un stimulus allant dans la direction préférentielle, les signaux inhibiteurs mettront tellement de temps à arriver que les signaux excitateurs auront déjà fait leur effet et déclenché une salve de potentiels d'action.
Les nerfs, chiasma et tractus optiques
[modifier | modifier le wikicode]Les axones des cellules ganglionnaires de la rétine se rassemblent pour former deux nerfs optiques, un par œil, qui sont des nerfs sensitifs qui partent de l’œil et innervent le cerveau. Mais les nerfs optiques ne se connectent pas directement sur le cerveau : ils font un nœud un peu avant d'entrer dans le cerveau. Pour être plus précis, les deux nerfs optiques se connectent au niveau de ce qu'on appelle le chiasma optique. Du chiasma optique sortent deux nerfs/faisceaux, appelés les tractus optiques, qui innervent directement le cerveau (le thalamus, pour être précis). Le tout est illustré ci-dessous.

La redistribution des axones dans le chiasma optique
[modifier | modifier le wikicode]
On pourrait croire que l’œil gauche envoie ce qu'il perçoit au cerveau droit et réciproquement pour le cerveau droit. C'est effectivement ce qui se passe chez certains animaux, comme le cheval ou les souris, mais pas chez les primates ou l'humain. Chez ces derniers, les axones se redistribuent en passant dans le chiasma optique, ce qui fait qu'il n'y a pas correspondance entre un nerf optique et un tractus optique. Tout dépend de l'animal et de la manière dont sont distribués les yeux sur le corps, ce qui nous amène à distinguer les animaux à vision latérale et frontale. Le schéma ci-contre illustre la différence entre animaux à vision frontale et à vision latérale.
Les animaux à vision dite latérale ont des yeux éloignés (la distance entre les deux yeux est "grande"), qui sont distribués chacun d'un côté du corps et qui regardent dans deux directions différentes (souvent opposées). Un bon exemple est la vision des poissons, qui ont un œil de chaque côté de leur corps, chaque œil regardant d'un côté du poisson. Par contre, les animaux à vision frontale ont les deux yeux qui pointent a peu-près vers la même direction et qui peuvent cibler le même point à regarder. Les yeux sont localisés sur leur tête, sur le visage et sont relativement proches, avec une faible distance inter-oculaire. Les champs visuels de chaque œil se recouvrent fortement avec une vision frontale, ce qui permet une vision binoculaire très précise, alors qu'ils se recouvrent peu avec une vision latérale et ne permettent pas de vision binoculaire notable. La vision latérale est surtout présente chez les proies, alors que les prédateurs ont surtout une vision frontale. La vision latérale permet en effet de couvrir un large champ de vision, certains poissons pouvoir carrément voir à 360 degrés autour d'eux, ce qui permet de plus facilement détecter les prédateurs. La vision frontale, quant à elle, permet de mieux voir un point précis, ce qui est utile pour observer une proie, ses mouvements, ses réactions, etc.
Chez les animaux à vision latérale, chaque hémisphère se connecte à un hémisphère et à lui seul. En général, la latéralisation est controlatérale : l’œil gauche se connecte à l'hémisphère droit et inversement. Cette organisation est parfaitement adaptée, car les deux hémisphères n'ont pas besoin de communiquer : chaque hémisphère traite son propre champ visuel indépendant de l'autre. La faible portion du champ visuel où la vision binoculaire est possible ne justifie pas une organisation plus complexe. De plus, les deux hémisphères peuvent communiquer par les faisceaux inter-hémisphériques, comme le corps calleux. Dans les grandes lignes, plus la portion binoculaire du champ visuel est grande, moins les animaux ont des hémisphères spécialisés sur le plan visuel. Le cas des animaux à vision frontale en sont un bon exemple : la portion binoculaire du champ visuel est clairement majoritaire, ce qui rend la spécialisation hémisphérique plus compliquée.

Chez les animaux à vision frontale, il y a redistribution des axones dans le chiasma optique. Mais dans les faits, les axones se redistribuent entre nerf optique droit et gauche dans le chiasma optique. Pour comprendre comment s’effectue cette redistribution, il faut faire une digression sur le champ visuel. Notre vision nous permet de voir dans un angle de plus de 180° autour de nous. On peut théoriquement diviser notre champ visuel en deux parties, à partir du milieu (du point de fixation, sur lequel on porte notre regard) : ces deux portions sont appelés des hémichamps visuels. Il faut prendre garde à ne pas confondre hémichamp et ce qui est perçu par l’œil gauche ou droit. Pour vous en rendre compte, fermez un œil : vous verrez que ce que vous voyez déborde quelque peu de l'hémichamp visuel correspondant. Avec la redistribution, le cerveau droit gère l'hémichamp gauche, et réciproquement pour le cerveau gauche. Pour preuve, il faut savoir que des lésions au chiasma optique entraînent parfois une cécité dans un hémichamp. Une telle perturbation est appelée une hémianopsie.
 |
 |
Les lésions du chiasma optique
[modifier | modifier le wikicode]Il arrive que le chiasma optique soit lésé ou comprimé, pour des raisons diverses. Si tel est le cas, la lésion entraîne des symptômes visuels assez marqués : une perte de la vision des deux côtés, ou d'un seul côté, parfois par une vision trouble, ou d'autres troubles de la vision. Les symptômes exprimés varient suivant la nature de la lésion. Tout dépend si la lésion coupe complètement le chiasma optique ou si la lésion est partielle. De même, les lésions longitudinales (qui suivent la direction avant-arrière) n'ont pas le même effet que les lésions latitudinales (d'une tempe à l'autre) qui coupent la lésion entre les nerfs et les tractus optiques).
Les lésions latitudinales coupent le chiasma optique en deux, en coupant les axones entre le nerf optique et le tractus optique. Les informations visuelles ne peuvent donc pas passer la barrière du chiasma optique et n'arrivent plus au cerveau. La communication visuelle étant complètement coupée, cela entraîne une cécité, qui est totale ou partielle. Évidemment, les sections complètes entraînent des lésions bien plus sévères que les lésions partielles. Les lésions latitudinales complètes entraînent une cécité totale, les sections partielles entraînant une cécité limitée à une plus faible portion du champ visuel, généralement limitée à un seul œil. À l'opposé, les lésions longitudinales coupent la redistribution des axones dans le chiasma optique. Chaque hémisphère ne reçoit plus que la vision d'un seul œil, ce qui entraîne des troubles plus limités. Il s'agit, en général, de troubles de la perception de la profondeur et/ou d'une vision trouble.
Diverses maladies ou traumatismes peuvent endommager le chiasma optique. Si la lésion fait suite à un AVC ou un traumatisme crânien, le chiasma optique fait alors partie d'un ensemble plus large de lésions. Mais il existe des lésions bien plus localisées : elles ne coupent presque que le chiasma optique et n'ont pas d'autres conséquences, si ce n'est quelques lésions limitées sur les structures avoisinantes). Elles sont causées par une tumeur hypothalamiques/hypophysaires. Il faut dire que le chiasma optique est localisé juste au-dessus du complexe hypothalamico-hypophysaire (un ensemble de glandes neuroendocrines), les deux étant littéralement collés l'un à l'autre. Si une tumeur a lieu dans cette structure, elle grossit et compresse le chiasma optique, causant des troubles de la vision. Si la tumeur grossit suffisamment ou très vite, elle peut couper complètement le chiasma optique (même si ce cas est assez rare et n'a lieu que pour des tumeurs avancées).
Les cibles du tractus optique : les voies secondaires
[modifier | modifier le wikicode]Au-delà du chiasma optique, les axones se distribuent dans deux nerfs qui font synapse sur plusieurs structures. La destination principale est le thalamus et plus précisément le corps genouillé latéral, qui sert de relai entre la rétine et le cortex visuel. On peut dire que la liaison (rétine-thalamus-cortex visuel) est la voie principale, car c'est elle qui est à l'origine de la vision proprement dite. Les autres voies visuelles servent surtout pour des réflexes liés à la pupille ou aux mouvements de la tête, ou encore à réguler les rythmes veille-sommeil en fonction de la lumière du Soleil. Elles ne jouent pas le moindre rôle dans le sentiment subjectif de la vision, elles ne permettent pas d'identifier des couleurs, de reconnaître des objets, d'analyser une scène quelconque, etc. Elles sont donc appelées les voies secondaires. Il y a en tout quatre voies secondaires :
- une vers le pretectum, qui se charge du réflexe de dilatation de la pupille en réponse à une lumière intense ;
- une vers des noyaux qui commandent les muscles cilaires qui déforment le cristallin : les noyaux d'Edinger-Westphal ;
- une qui va vers le noyau-suprachiasmatique de l'hypothalamus et qui sert à réguler les rythmes veille-sommeil selon la luminosité ;
- une autre vers le colliculus supérieur, qui se charge de mouvements réflexes de la tête en réponse à des sensations visuelles.
Dans ce qui va suivre, nous allons parler des voies secondaires, la voie principale étant laissée à plus tard.
Le Pretectum et les noyaux d'Edinger-Westphal
[modifier | modifier le wikicode]Les noyaux d'Edinger-Westphal commandent les muscles de l'iris et du cristallin. Les muscles de l'iris sont ceux qui dilatent ou contractent la pupille. Quand la pupille est contractée, il s'agit d'une myosis, alors que la dilation est appelée mydriase. D'ordinaire, ces deux phénomènes ont lieu suite à une variation de luminosité. Une mydriase ou un myosis non-réactive, c'est à dire qui ne disparaît pas quel que soit les circonstances, est un signe de souffrance cérébrale. Elle est souvent signe d'une hypertension intracrânienne ou d'une surdose de médicaments/drogues. Par exemple, un myosis peut être causé par une intoxication aux opiacés, ou aux inhibiteurs de l'acétylcholinéstèrase (certains médicaments, gaz de combat de type sarin ou VX, ...). Par exemple, quelqu'un qui fait une overdose de morphine ou d’héroïne exprimera un myosis manifeste.

L'iris se contracte quand la lumière est trop importante, histoire de limiter la quantité de lumière captée par la rétine. Le réflexe inverse a lieu quand la luminosité baisse. Ce réflexe est commandé par les noyaux d'Edinger-Westphal, avec cependant l'intervention du pretectum. Dans les grandes lignes, les récepteurs envoient des informations sur la luminosité au pretectum, qui mesure la luminosité et commande les noyaux d'Edinger-Westphal. Ceux-ci vont alors dilater ou contracter la pupille. Il faut signaler que ce réflexe touche les deux yeux en même temps : la pupille de l’œil gauche se contracte en même temps que celle de l’œil droit, avec la même intensité. Si ce n'est pas le cas, c'est signe de souffrance cérébrale importante, provenant d'une hémorragie cérébrale ou d'un AVC.

Autre réflexe pris en charge par les noyaux d'Edinger-Westphal : l’accommodation. Celle-ci consiste à modifier la forme du cristallin, de manière à ce que les rayons lumineux se concentrent sur la rétine. Ce cristallin sert en quelque sorte de lentille, pour les yeux. Il faut signaler que la pupille peut se contracter ou se dilater suite à l’accommodation. Ce réflexe est intégralement commandé par les noyaux d'Edinger-Westphal, sans intervention du pretectum. Si le pretectum est lésé, alors que les noyaux d'Edinger-Westphal sont intacts, on observe une suppression de la contraction/dilation pupillaire lors de variation de luminosité, mais celle-ci reste présente quand le cerveau accommode. C'est ce qu'on appelle le signe d'Argyll Robertson.
Les noyaux supra-chiasmatiques
[modifier | modifier le wikicode]Juste au-dessus du chiasma optique, on trouve un noyau hypothalamique innervé par le nerf optique : le noyau supra-chiasmatique (littéralement, au-dessus du chiasma). Les cellules ganglionnaires qui font synapse avec ce noyau sont les cellules ganglionnaires photosensibles (celles qui contiennent du pigment, la mélanopsine, qui s'active en réponse à l'intensité lumineuse). Ainsi, l'hypothalamus est informé de l'intensité lumineuse perçue par les yeux et peut en déduire s'il fait jour ou nuit. Ce faisant, il envoie des commandes à la glande pinéale, une structure de l'épithalamus qui secrète un somnifère naturel : la mélatonine. C'est ainsi que l'on devient somnolent ou que l'on commence à s'endormir : c'est parce que la rétine a informé une portion de l'hypothalamus (les noyaux suprachiasmatiques) que le jour est tombé et qu'il faudrait faire produire de la mélatonine à la glande pinéale. Nous parlerons plus en détail de ce noyau dans le chapitre sur les rythmes circadiens.
Le colliculus supérieur
[modifier | modifier le wikicode]Une dernière voie-non-corticale fait synapse sur le colliculus supérieur. Pour faire simple, celui-ci se charge de l'orientation du regard vers les objets à fixer. Il commande les mouvements de la tête et les mouvements rapides des yeux. Dans le détail, il commande ce qu'on appelle les saccades oculaires, des mouvements rapides des yeux qui permettent de fixer ceux-ci sur une nouvelle cible. Nous reparlerons de tout cela dans le chapitre sur la motricité oculaire. Tout ce que nous pouvons dire pour le moment est que cette fonction est redondante avec une bonne partie du cortex moteur, qui commande elle aussi les saccades oculaires. La raison est que le colliculus est plus ancien dans l'évolution, les aires corticales oculomotrices étant présentes uniquement chez les mammifères. Chez les vertébrés "inférieurs", seul le colliculus est en charge des saccades oculaires. Par la suite, les mammifères ont vu croître le cortex moteur, qui a pris en charge les saccades oculaires.
On pourrait croire qu'il s'agit d'un noyau purement moteur, mais ce n'est pas le cas. Il intègre les informations provenant du nerf optique et détermine où déplacer le regard, ce qui en fait un noyau à la fois moteur et sensoriel/visuel. Cette double fonction d'analyse visuelle et de commande motrice se voit dans son organisation anatomique. Le colliculus supérieur est composé de plusieurs couches de neurones, regroupées en deux sections : les couches visuelles (stratum opticum) et les couches motrices. Les couches les plus en surface sont des couches dites optiques, ce nom trahissant le fait qu'elles sont innervées par le nerf optique et effectuent des traitements visuels. Les couches plus profondes sont purement motrices et commandent le mouvement des yeux. Le nombre de couches dépend de l'animal considéré et il est de 7 couches chez l'humain : approximativement 3 couches visuelles et 4 couches motrices.
Les cibles du tractus optique : la voie principale
[modifier | modifier le wikicode]Après avoir vu les voies secondaires, il est temps de voir la voie principale, qui relie la rétine au corps genouillé latéral, par l'intermédiaire du tractus optique. La destination principale du tractus optique est le thalamus et plus précisément le corps genouillé latéral, qui sert de relai entre la rétine et le cortex visuel. Si le cortex visuel est le lieu de traitement final de l'information visuelle, le corps genouillé latéral est un simple relai qui ne semble pas faire beaucoup de traitements visuels. L'information transmise par la rétine ressort du thalamus sans avoir été modifiée de manière substantielle.
Le corps genouillé latéral du thalamus
[modifier | modifier le wikicode]

Les corps genouillés sont composés de six couches de neurones. Le corps genouillé latéral est, dans cette voie, un simple relai qui ne semble pas faire beaucoup de traitements visuels. L'information transmise par la rétine ressort du thalamus sans avoir été modifiée de manière substantielle. Les deux demi-rétines gauches se projettent sur le corps genouillé latéral de l'hémisphère gauche, alors que les deux demi-rétines droites se connectent sur le corps genouillé latéral de l'hémisphère droit. Mais il faut noter que la connexion de chaque hémi-champ visuel aux corps genouillés latéraux est assez particulière. Le corps genouillé latéral est composé de plusieurs couches, dont le nombre dépend de l'espèce. Les axones de l'hémi-champ visuel gauche font synapse sur certaines couches, alors que ceux liés à l'hémi-champ droit vont se connecter sur d'autres couches.
Chez le chat, il n'y a que trois couches : une couche supérieure et une couche inférieure composées de grosses cellules et une couche mineure de petites cellules. La couche supérieure est la plus épaisse et surmonte la couche inférieure, plus fine. La couche supérieure est connectée à l’œil controlatéral, alors que la couche inférieure est connectée à l’œil ipsilatéral.
Chez l'homme, on trouve 6 couches différentes posées les unes sur les autres et séparées par des lamelles de substance blanche. La rétine de l’œil controlatéral se connecte aux couches 1, 4 et 6 (œil gauche pour le thalamus droit et réciproquement), tandis que la rétine de l'œil ipsilatéral fait synapse sur les couches 2,3 et 5. Par exemple, dans le corps genouillé gauche, l’œil gauche se connecte aux couches 1, 4 et 6, alors que l’œil droit se connecte aux couches 2, 3 et 5. Et c'est l'inverse dans le corps genouillé droit. Précisons cependant que cette structuration ne se trouve pas chez les animaux avec un système visuel simple : on ne l'observe pas chez les souris, par exemple.

Dans le corps genouillé latéral, on retrouve la différence entre voie magnocellulaire et parvocellulaire. Les cellules ganglionnaires qui se connectent sur les couches 1 et 2 sont différentes de celles qui se connectent sur les autres couches. Les couches 1 et 2 correspondent à la voie magnocellulaire, alors que les autres couches correspondent respectivement à la voie parvocellulaire et à une autre voie dite koniocellulaire, dont les neurones sont situés entre les couches 4 et 6. Les lésions dans les couches 1 et 2 ne diminuent pas la capacité de reconnaître les couleurs mais perturbent la capacité de détecter les objets qui se déplacent, alors que les lésions dans les autres couches ont l'effet exactement inverse.
La radiation optique
[modifier | modifier le wikicode]
Le corps genouillé latéral émet un faisceau appelé la radiation optique, qui fait synapse dans le lobe occipital du cortex. Elle a une forme légèrement recourbée, en forme de virgule. Il se connecte au cortex occipital ipsilatéral, dans le même hémisphère que le corps genouillé latéral émetteur, ce qui fait qu'il n'y a pas de radiation optique inter-hémisphérique. On peut voir que chaque radiation optique (il y en a deux, une par hémisphère), est composée de plusieurs sous-faisceaux indépendants, dont les trois principaux portent les noms de faisceau dorsal, de faisceau central/ventral et de boucle de Meyer.
 |
 |
Pour faire simple, on peut subdiviser chaque radiation optique en deux sections : la boucle de Meyer et les autres faisceaux (regroupés dans la boucle de Baum). La boucle de Meyer forme la partie inférieure de la radiation optique, alors que le reste forme sa subdivision supérieure. Cette distinction a une importance clinique assez importante. Si chaque radiation optique transmet la vision dans un hémi-champ visuel, les subdivisions inférieure et supérieure prennent en charge chacune une moitié de l'hémi-champ visuel. La portion inférieure transmet la vision sur la moitié haute de l'hémi-champ visuel, alors que portion supérieure transmet la vision dans sa partie basse. Pour résumer, on peut découper le champ de vision total en deux hémi-champ, chacun découpé quatre quadrants : un quadrant supérieur pour la portion haute de l'hémi-champ et un quadrant inférieur pour la portion basse. Les subdivisions supérieures et inférieures de la radiation optique transmettent respectivement la vision dans le quadrant inférieur et supérieur (attention à l'inversion).
Une lésion d'une seule radiation optique entraîne une hémianopsie (cécité dans un hémi-champ visuel) ipsilatérale. Les lésions bilatérales, qui coupent les deux radiations optiques, entraînent une cécité. En général, la cécité se limite à un quadrant, à savoir à la moitié haute ou basse d'un hémi-champ visuel, ce qui fait qu'on parle d'hémianopsie en quadrant, ou encore de quadranopsie. Les lésions du faisceau de Meyer suppriment la vision dans le quadrant haut, alors que les lésions des deux autres faisceaux causent une cécité dans le quadrant bas.
Le cortex visuel
[modifier | modifier le wikicode]Une fois que les informations visuelles sont passées dans le thalamus, elles sont envoyées dans le lobe occipital, siège du cortex visuel. Celui-ci est découpé par les scientifiques en deux grandes parties : les aires de bas niveau et les aires de haut niveau. Les aires primaires, aussi appelées aires de bas niveau, traitent les informations purement perceptives des stimulus : couleur, luminosité, etc. Elles permettent de reconnaître les formes ou les objets connus, éventuellement de trouver des informations spatiales, sur le mouvement des objets. Les aires du cortex de haut niveau, aussi appelé cortex secondaire, traitent le sens de ce qui est perçu : ces aires comparent ce qui est vu avec des informations conceptuelles en mémoire, ce qui permet de classer ou de nommer les objets perçus.

Le cortex visuel primaire
[modifier | modifier le wikicode]
Après avoir fait relais dans les corps genouillés du thalamus, les informations visuelles arrivent dans le néocortex visuel primaire. Si les neurones de la couche 4 du cortex primaire sont spécifiques à l’œil gauche ou droit, ce n'est pas le cas des autres couches : hormis la couche 4, la ségrégation entre informations perçues par l’œil droit et l’œil gauche disparaît.
-
 Cortex strie.
Cortex strie. -
 Scissure du cortex strié et place des couches V3, V2 et V1.
Scissure du cortex strié et place des couches V3, V2 et V1.


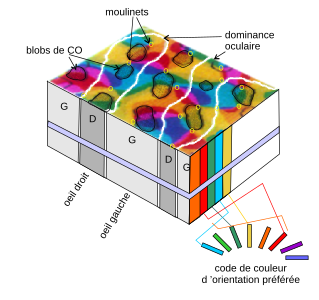
Si on plonge une électrode à la verticale dans le cortex visuel primaire à des profondeurs différentes, on s'aperçoit que toutes les cellules rencontrées sur le chemin de l’électrode émettent des potentiels d'action quand une stimulation bien précise est présente sur un champ récepteur qui leur est dédié. Par exemple, elles émettent quand le champ récepteur voit un objet présenté avec un certain angle. Et ce qui est valable pour l'orientation est aussi valable pour la couleur ou la distance de l'objet présenté. Bref, il semblerait que le cerveau soit organisé en colonnes corticales, des colonnes de neurones qui répondent à des stimulations identiques bien précises, stimulations qui doivent être présentées sur un champ récepteur bien précis de la rétine. On trouverait:
- des colonnes spécialement dédiées à la détection d'un objet présenté suivant un angle précis : les colonnes d'orientation ;
- d'autres spécialisées dans la couleur : les colonnes de couleur ;
- d'autres dans la perception de la profondeur : les colonnes de dominance oculaire ;
- d'autres s'activent quand un stimulus est présenté sur un œil précis, mais pas sur l'autre ;
- etc.
Des colonnes corticales proches ont des champs récepteurs qui sont identiques ou très proches sur la rétine : ces colonnes immédiatement adjacentes ont des champs récepteurs qui se recouvrent. Si on oriente une électrode perpendiculairement à la surface du cortex, on s’aperçoit rapidement que des colonnes d'orientation adjacentes perçoivent des angles similaires : des colonnes adjacentes codent des angles qui varient de 10° en 10° dans le même champ récepteur. Le même chose a lieu pour les autres types de colonnes. L'ensemble forme une organisation très complexe, encore mal comprise à l'heure actuelle.
-
 Colonnes corticales d'orientation dans la couche V1 du cortex visuel.
Colonnes corticales d'orientation dans la couche V1 du cortex visuel. -
 Cartes superposées.
Cartes superposées. -
 Hypercolonne.
Hypercolonne.



Le cortex visuel secondaire
[modifier | modifier le wikicode]Au-delà du cortex primaire, on sait que les informations visuelles progressent dans une série d'aires cérébrales dédiées à la vision. Mais ces aires sont relativement mal identifiées. Tout ce que l'on sait, c'est qu'elles sont localisées aussi bien dans les lobes occipitaux que dans les lobes pariétaux et temporaux. Dans les grandes lignes, les chercheurs ont identifié deux voies de traitement des informations visuelles :
- une voie ventrale, spécialisée dans la reconnaissance des objets et des couleurs, qui envoie des axones vers le lobe temporal ;
- une voie dorsale, spécialisée dans le mouvement qui envoie des informations vers le cortex pariétal.

Une première preuve de cette distinction provient de l'étude des lésions des cortex temporaux et pariétaux. Les deux se traduisent par des déficits très différents. Des lésions dans le cortex pariétal causent des problèmes dans la cognition spatiale, tandis que des lésions dans le cortex temporal entraînent des problèmes dans la reconnaissance des objets.
La voie dorsale : l'analyse de la position et du mouvement
[modifier | modifier le wikicode]Les lésions dans le cortex pariétal se traduisent par des déficits spatiaux, dans une certaine mesure. Les patients au cortex pariétal lésé ne peuvent plus percevoir le mouvement, alors qu'ils peuvent reconnaître les couleurs et les objets et que leur capacité à faire des distinctions visuelles fines est intacte. Ils ne perçoivent plus le mouvement, au point que leur vision ressemble à une suite de diapositives. Ce trouble est appelé l'akinétopsie. Cependant, tout le cortex pariétal n'est pas impliqué dans la cognition spatiale, mais seulement une partie, le reste étant dédié à la perception tactile. Rappelons que le cortex pariétal est subdivisé par le sillon intrapariétal en plusieurs gyrus, nommés gyrus postcentral, gyrus supérieurs et inférieurs. Le gyrus supérieur et le gyrus postcentral sont dédiés à la perception tactile, alors que le gyrus inférieur est dédié à la cognition spatiale.
La voie ventrale : reconnaissance des objets
[modifier | modifier le wikicode]
La voie ventrale est composée du cortex temporal inférieur, qui regroupe les deux gyrus temporaux inférieurs et médian. On sait que celui-ci intervient dans la reconnaissance des objets grâce aux études de lésions. Des lésions dans cette région causent des déficits dans la capacité à reconnaître, dessiner et apparier des informations visuelles. Les premières études sur le sujet furent celles de Kluver et Bucy, datées de 1938. L'étude princeps étudiait l'ablation du cortex temporal sur des macaques. Il est rapidement apparu que cette lésion entraînait une constellation de symptômes divers : sexualité incontrôlée, docilité, tentative de consommation d'objets qui ne peuvent pas être mangés, abolition de la peur, troubles de la mémoire et troubles de la reconnaissance visuelle. Cette constellation de symptômes est aujourd'hui appelée syndrome de Kluver-Bucy. Il est rapidement apparu que seuls les troubles de la reconnaissance visuelle provenait de l'ablation du cortex inférotemporal, les autres troubles s'expliquant par l'ablation de l'amygdale, de l'hippocampe et du reste du cortex temporal.
Chez certains patients, la capacité de reconnaître des couleurs est dégradée à la suite d'une lésion cérébrale. Certains voient carrément la vie en noir et blanc, avec des niveaux de gris qui correspondent à la luminosité de ce qu'ils regardent : la couleur a disparu. En revanche, ces patients n'ont aucun déficit pour percevoir des objets en mouvement : ils arrivent notamment à distinguer des objets qui bougent rapidement d'objets lents. On parle d'achromatopsie cérébrale. Généralement, les lésions qui causent ce trouble sont localisées dans le cortex temporal, avec une possible préférence pour le cortex inférotemporal.
D'autres patients sont incapables de reconnaître les objets qui leur sont présentés, de les nommer ou de les classer : ils sont atteints d'agnosie. Lorsqu'on leur présente un objet ou un visage, ces patients sont incapables de le nommer, de le classer ou de donner des informations pertinentes à son propos. Techniquement, l'agnosie peut toucher toute modalité sensorielle, mais nous allons nous concentrer sur les agnosies visuelles. Grossièrement, on peut classer les agnosies en deux types selon leur cause : les agnosies perceptives et associatives. Les premières sont causées par un déficit perceptif, le patient ne pouvant plus percevoir visuellement les objets. En comparaison, les agnosies associatives proviennent d'une incapacité à attribuer un sens à ce qu'ils voient : les patients ne peuvent pas associer ce qu'ils voient avec les représentations conceptuelles de ces objets en mémoire. La différence entre ces deux agnosies est assez facile à mettre en évidence : les patients avec une agnosie perceptive ne peuvent pas recopier un dessin, une photographie ou une image qu'on leur présente, alors que ceux atteint d'une agnosie associative le peuvent. Certaines formes d'agnosies touchent des stimulus précis, mais pas l'intégralité des objets : les patients n'ont des déficits que pour une catégorie limitée d'objets ou de stimulus. En voici quelques exemples :
- Certaines agnosies ne touchent que l'écriture. Les patients atteints d'alexie pure ne peuvent plus lire : ils ne peuvent plus reconnaître les lettres ou les mots.
- D'autres sont incapables de nommer des couleurs, cette agnosie des couleurs étant cependant assez rare.
- Plus rarement, certains patients ont des déficits qui se limitent à la reconnaissance des lieux : on parle alors d'agnosie environnementale.
- Certaines agnosies dégradent la capacité de reconnaître les visages : on parle alors de prosopagnosie. Les patients atteints de ce trouble ne peuvent pas reconnaître les visages de leurs proches, amis ou connaissances. Ils peuvent voir les visages, les décrire, et n'ont pas de déficits de perception : il s'agit donc d'agnosies associatives. Ils peuvent parfois identifier le sexe ou l'âge de la personne quand on leur présente un visage (sur photographie, ou en personne), preuve que certaines capacités perceptives et associatives sont conservées. Les prosopagnosiques peuvent cependant reconnaître les voix des personnes avec qui ils conversent, ce qui leur permet d'identifier leur interlocuteur, preuve que la prosopagnosie est un déficit de la reconnaissance visuelle.
L'origine de ces syndromes s'expliquerait par la localisation des lésions à l'origine des déficits. Par exemple, la prosopagnosie proviendrait de lésions qui ne sont pas à la même place que celles qui causent une alexie. Ainsi, le cerveau aurait une aire cérébrale dédiée à la reconnaissance des visages (fusiform face area), une autre aire pour les mots (la visual word form area), une autre pour les lieux (parrahippocampal place area) et une autre pour les formes et/ou les couleurs. Les preuves de l'existence de telles aires spécialisées sont nombreuses. Dans les études d'imagerie, on voit clairement que chaque aire s'active quand on présente un type de stimulus précis (un visage pour le gyrus fusiforme, ou un mot dans la word form area), preuve que ces aires sont spécialisées. L'étude des lésions chez l'humain et le macaque va aussi dans le même sens. Enfin, certaines études ont étudié l'activité électrique de neurones uniques : celles-ci ont montré que certains neurones ne répondaient qu'à des visages ou qu'à des objets ou lieux.
.png)