
Systèmes sensoriels/Système Visuel
Introduction
Système auditif
Système vestibulaire
Système somatosensoriel
Système olfactif
Système gustatif
Introduction
[modifier | modifier le wikicode]De manière générale, les systèmes visuels s'appuient sur les ondes électro-magnétiques (EM) pour donner à un organisme plus d'informations sur son environnement. Ces informations pourraient porter sur les partenaires potentiels, les dangers et les sources de subsistance. Différents organismes ont différents constituants qui composent ce que l'on appelle un système visuel.
La complexité des yeux va du simple ocelle, qui n'est rien de plus qu'une collection de cellules photosensibles, à un « objectif d'appareil photographique » à part entière. Un organisme qui possède différents types de cellules photosensibles ou de cellules sensibles à différentes gammes de longueurs d'onde est théoriquement capable de percevoir diverses couleurs ou au moins des différences de couleur.
La polarisation, une autre propriété du rayonnement électromagnétique, peut être détectée par certains organismes, les insectes et les céphalopodes ayant la plus grande précision.
Dans ce texte, l'accent a été mis sur l'utilisation des ondes EM dans la vision. Certes, certains organismes ont mis au point d'autres moyens de voir ou d'ajouter à ce qu'ils voient des informations extra-sensorielles, comme les baleines ou les chauves-souris qui utilisent l'écho-localisation pour percevoir leur environnement, mais nous choisirons d'exclure ces techniques de notre étude. En effet, vision et visuel sont les mots les plus souvent associés aux ondes électromagnétiques de la gamme des longueurs d'onde visuelles, qui est habituellement définie comme celle de la vision humaine. Étant donné que certains organismes détectent les ondes électromagnétiques avec des fréquences inférieures et supérieures à celles de l'homme, une meilleure définition doit être établie.
Nous choisirons ici de définir la gamme de longueurs d'onde visuelles comme celles qui sont comprises entre 300 nm et 800 nm. Cela peut sembler arbitraire mais nous pouvons ainsi inclure la vision de certains oiseaux dans le spectre choisi. De plus, avec cette gamme de longueurs d'onde, nous excluons la vision thermique de certains organismes comme les serpents. En effet, les serpents utilisent leurs fossettes sensorielles, qui sont sensibles aux émissions électromagnétiques entre 5 000 et 30 000 nm, donc dans l'infrarouge ; cependant, même s'ils perçoivent de loin ces rayonnements, ils ne les « voient » pas en ce sens qu'il n'y a aucune formation d'image. Cependant, des spécimens aveugles, capables de viser et d'attaquer des parties du corps particulières ont été documentés.
Dans un premier temps, une brève description des différents types d'organes sensoriels du système visuel sera développée, suivie d'une explication approfondie des composants de la vision humaine et du traitement du signal visuel chez l'homme. Puis nous finirons par un exemple des possibilités de perception.
Organes sensoriels
[modifier | modifier le wikicode]La vision, ou capacité de voir, dépend des organes sensoriels du système visuel, les yeux. Les schémas de constitution des yeux sont divers et variés. On les classe selon leur complexité, elle-même dépendante des besoins de l'organisme. Ces différentes constitutions ont des capacités différentes, sont sensibles à différentes longueurs d'onde et présentent différents degrés d'acuité. Elles nécessitent également des traitements différents pour interpréter les données et des paramètres différents pour fonctionner de manière optimale. La capacité de détecter et de déchiffrer les ondes EM est devenue être un atout précieux pour la plupart des formes de vie, augmentant les chances de survie des organismes qui les utilisent. Dans des environnements sans lumière suffisante ou présentant une absence totale de lumière, les formes de vie possédant la vue n'en retirent aucun avantage particulier, ce qui a finalement entraîné une atrophie des organes sensoriels visuels et une dépendance accrue des autres sens. C'est le cas notamment pour certains animaux des cavernes, les chauves-souris, etc. Il est intéressant de noter que les organes sensoriels visuels sont ajustés sur la fenêtre optique, définie comme étant les longueurs d'onde EM entre 300 nm et 1100 nm qui traversent l'atmosphère pour atteindre le sol. Ceci est montré dans la figure ci-dessous. Vous remarquerez peut-être qu'il existe d'autres « fenêtres », une fenêtre infrarouge (IR), qui explique dans une certaine mesure la "vision" thermique des serpents et une fenêtre dans les radiofréquences (RF), qu'aucune forme de vie connue n'est capable de détecter.
Au fil du temps, l'évolution a donné lieu à de nombreux schémas de constitution oculaires qui, à la suite d'évolutions successives, ont amené certains organismes similaires à partager les mêmes spécificités. Indépendamment de l'espèce et de la complexité du type d'organe sensoriel, il existe un élément essentiel que l'on retrouve universellement : l'utilisation des protéines sensibles à la lumière, les opsines. Sans trop se concentrer sur la base moléculaire, les différents schémas de construction peuvent être classés comme suit :
• Yeux simples ou camérulaires : ocelle, œil en trou d'épingle, œil à cristallin sphérique, œil miroir...
• Yeux composés.
La configuration la plus simple des yeux ne permet aux organismes que de détecter la lumière ambiante et donc de savoir s'il y a ou non de la lumière. Il s'agit normalement d'un simple rassemblement de cellules photosensibles formant un groupe, au même endroit, ce que l'on appelle parfois une « tache oculaire » ou stemma. Grâce aux mouvements des yeux, les organismes ont accès aux informations directionnelles, condition essentielle à la formation d'images. Ces fossettes sensorielles sont de loin les types d'organes sensoriels visuels les plus communs et on peut les trouver dans plus de 95 % des espèces connues.
.jpg)
En suivant cette approche jusqu'à l'extrême, on peut considérer que la fossette sensorielle est une simple structure caverneuse qui augmente l'acuité de l'image, contre une perte d'intensité. En d'autres termes, il existe un compromis entre intensité, ou luminosité, et acuité. Considérons maintenant le Nautilus, espèce appartenant à la famille des Nautilidae, organismes considérés comme des « fossiles vivants ». Ce sont les seules espèces connues qui possèdent l’œil à sténopé, comparable à la camera obscura. Tout comme les appareils photo, les Nautilus sont capables d'ajuster la taille de l'ouverture, augmentant ou diminuant la résolution de l’œil lorsque la luminosité de l'image diminue ou augmente. De même, pour régler le compromis intensité-résolution, il convient d'ajouter un objectif, c'est-à-dire une structure qui dirige la lumière vers une zone centrale, celle qui, le plus souvent, contient une plus grande densité de capteurs photoélectriques. En ajustant la forme du cristallin et en la déplaçant, ainsi qu'en contrôlant la taille de l'ouverture de la pupille, les organismes peuvent s'adapter à différentes conditions et fixer leur attention sur des zones d'intérêt particulières. La dernière amélioration apportée aux divers schémas de constitution oculaires déjà mentionnés est l'inclusion d'une cornée réfractrice. Les deux tiers de la capacité visuelle de ce type d’œil résident dans l'utilisation d'un liquide à indice de réfraction élevé situé à l'intérieur de la cornée, permettant une vision à très haute résolution. Ce schéma est celui des yeux des humains et de la plupart des animaux. Au sein d'un même catégorie d'yeux, il existe de nombreuses variations de la structure du cristallin, du nombre de cristallins, de la densité du photocapteur, de la forme de la fovéa, du nombre de fovéas, de la forme de la pupille, etc. afin d'augmenter les chances de survie de l'organisme en question. Ces variations engendrent diverses apparences externes des yeux, au sein d'une même catégorie, comme le montre la série de photographies d'animaux appartenant à la même catégorie d'yeux (à cornée réfractrice) présentée ci-dessous.




.jpg)
Une autre alternative est celle des yeux des mollusques. Au lieu de focaliser la lumière de manière conventionnelle en un seul point à l'arrière de l’œil, grâce à un cristallin ou à un système de cristallins, ces organismes ont à l'intérieur de la chambre oculaire des structures semblables à un miroir qui réfléchissent la lumière dans une partie centrale. Bien qu'il n'existe pas d'exemples d'organismes dont les yeux réfléchissants soient capables de former une image, il existe une espèce de poissons, les revenants (Opisthoproctidae), qui les utilise en combinaison avec des yeux à cristallins.

Le dernier type correspond aux yeux composés des insectes et des crustacés. Ces yeux sont constitués d'un certain nombre de sous-unités fonctionnelles appelées ommatidies, dont chacune est composée d'une facette, ou surface frontale, d'un cône cristallin transparent et de cellules photosensibles. En outre chacune des ommatidies est isolée par des cellules pigmentaires, ce qui garantit que la lumière incidente qu'elle reçoit est formée de rayons aussi aussi parallèles que possible.
La combinaison des sorties de chacune de ces ommatidies forme une image en mosaïque, avec une résolution proportionnelle au nombre d'unités d'ommatidies. Par exemple, si les humains avaient des yeux composés, ceux-ci recouvriraient tout leur visage pour conserver la même résolution. Il convient de noter qu'il existe de nombreux types d'yeux composés différents, mais approfondir ce sujet dépasse le cadre de ce texte.

Chez un animal on peut trouver des yeux de diverses constitution et en nombre variable selon l'espèce. Si les humains disposent de deux yeux, les araignées en possèdent généralement huit, dont la taille dépend des fonctions qui leur sont assignées. Les araignées sauteuses, par exemple, ont deux grands yeux frontaux qui leur donnent une excellente acuité visuelle, utile pour repérer et suivre leurs proies, et six yeux plus petits dont la résolution beaucoup plus faible leur permet de détecter les mouvements qui surviennent dans leur environnement. Deux photographies des yeux d'une araignée sauteuse et d'une araignée-loup illustrent la variabilité de la disposition des yeux des arachnides. Beaucoup d'insectes disposent à la fois d'yeux composés et d'ocelles ; les yeux composés des libellules présentent des structures différentes vers le haut et vers le bas.

Anatomie du système visuel
[modifier | modifier le wikicode]
Nous, êtres humains, sommes des créatures visuelles. Par conséquent, nos yeux sont compliqués et dotés de nombreux composants. Dans ce chapitre, nous essayons de décrire ces composants, donnant ainsi un aperçu des propriétés et des fonctionnalités de la vision humaine.
À l'intérieur du globe oculaire : pupille, iris et cristallin
[modifier | modifier le wikicode]Le flux lumineux qui pénètre dans l’œil par la pupille située à l'avant dépend directement des dimensions de cette ouverture, un « trou » dont l'aspect noir est dû à l'absorption presque totale de la lumière par les tissus intérieurs. L'iris qui entoure la pupille humaine peut être diversement coloré par des pigments.
Outre cette couche de pigments, l'iris possède deux couches de muscles ciliaires : le sphincter pupillaire se contracte pour rendre la pupille plus petite tandis que le dilatateur pupillaire se contracte pour la dilater. La combinaison de ces muscles antagonistes peut ainsi d'ouvrir plus ou moins la pupille en fonction des besoins ou des conditions de la personne. Les muscles ciliaires sont commandés par des zonules ciliaires, des fibres qui modifient également la forme du cristallin et le maintiennent en place.
L'étude comparée de l'iris et du diaphragme d'un objectif photographique fait apparaître de notables différences.
Le cristallin est situé immédiatement derrière la pupille. Sa forme et ses caractéristiques sont similaires à ceux des objectifs d'appareil photo mais il fonctionne différemment. La forme du cristallin est ajustée par la traction des zonules ciliaires, ce qui modifie par conséquent la distance focale. Avec l'aide de la cornée, le cristallin peut modifier la focalisation, une spécificité qui le rend essentiel au fonctionnement de l’œil, même si seulement un tiers de la capacité visuelle est permise par le cristallin lui-même. Il est aussi le filtre principal de l’œil. La majeure partie du cristallin est formée de fibres cristallines : des cellules longues et fines, dépourvues de la plupart des mécanismes cellulaires afin de favoriser la transparence. Avec les protéines hydrosolubles appelées cristallines, elles augmentent l'indice de réfraction du cristallin. Les fibres jouent également un rôle dans la structure et la forme du cristallin elles-mêmes.
La formation des images dans l’œil. La cornée et son agent protecteur, la sclérotique
[modifier | modifier le wikicode]La cornée, responsable des deux tiers restants de la capacité visuelle, recouvre l'iris, la pupille et le cristallin. Elle concentre les rayons lumineux avant qu'ils traversent l'iris avant et le cristallin. La cornée n'a que 0,5 mm d'épaisseur et se compose de 5 couches:
• Épithélium : couche de tissu épithélial recouvrant la surface de la cornée.
• Membrane de Bowman : épaisse couche protectrice composée de fibres de collagène solides qui maintiennent la forme générale de la cornée.
• Stroma : couche composée de fibrilles de collagène parallèles et représentant 90 % de l'épaisseur de la cornée.
• Membrane de Descemet et endothélium : deux couches ajustées à la chambre antérieure de l’œil, remplies d'un fluide produit par le corps ciliaire, l'humeur aqueuse. Ce fluide hydrate le cristallin, le nettoie et maintient la pression dans le globe oculaire. La chambre située entre la cornée et l'iris contient un corps trabéculaire, à l'intérieur duquel le liquide est drainé par le canal de Schlemm, à travers la chambre postérieure.
La surface de la cornée est située sous deux membranes protectrices appelées sclérotique et capsule de Tenon. Ces deux couches protectrices enveloppent complètement le globe oculaire. La sclérotique est constituée de fibres élastiques de collagène qui protègent l’œil des agressions extérieures. Cette couche forme le blanc de l’œil. Elle est traversée de nerfs et de vaisseaux, le plus grand étant le nerf optique. Elle est recouverte par la conjonctive, qui est une membrane muqueuse claire à la surface du globe oculaire. Cette membrane tapisse également l'intérieur de la paupière. Elle est lubrifiée et protégée par les larmes produites par les glandes lacrymales. La couche protectrice restante, la paupière, sert également à répandre ce lubrifiant autour du globe oculaire.

Faire bouger les yeux. Les muscles extra-oculaires
[modifier | modifier le wikicode]Le globe oculaire se déplace grâce à une structure musculaire compliquée comprenant quatre muscles droits : inférieur, médial, latéral et supérieur et deux muscles obliques : inférieur et supérieur. Le positionnement de ces muscles est présenté ci-dessous, avec les fonctions suivantes :
Comme vous pouvez le constater, les muscles extra-oculaires (2,3,4,5,6,8) sont attachés à la sclérotique du globe oculaire et prennent naissance dans l'anneau de Zinn, un tendon fibreux entourant le nerf optique. Un système de poulie est créé avec la trochlée (la trochlée étant la poulie et le muscle oblique supérieur étant la corde), permettant d'orienter correctement la force musculaire. Les muscles extra-oculaires restants ont un chemin direct vers l’œil et ne nécessitent donc pas de systèmes de poulies. En utilisant ces muscles extra-oculaires, l’œil peut pivoter vers le haut, le bas, la gauche, la droite et des mouvements alternatifs sont possibles.
Les mouvements sont également très importants pour que nous puissions voir. Les mouvements de vergence permettent le bon fonctionnement de la vision binoculaire. Les mouvements rapides inconscients, appelés saccades, sont essentiels pour que les personnes puissent garder la mise au point sur un objet. La saccade est une sorte de mouvement nerveux effectué lorsque les yeux balayent le champ visuel afin de déplacer légèrement le point de fixation. Lorsque vous suivez un objet en mouvement avec votre regard, vos yeux exécutent ce que l'on appelle une poursuite visuelle. Des mouvements involontaires supplémentaires, appelés nystagmus, sont causés par les signaux du système vestibulaire. Ensemble, ils forment les réflexes vestibulo-oculaires.
Le tronc cérébral contrôle tous les mouvements des yeux, différentes zones étant responsables de différents mouvements.
• Pons : mouvements horizontaux rapides, tels que saccades ou nystagmus
• Mésencéphale : mouvements verticaux et de torsion
• Cervelet : réglage précis
• Noyau Edinger-Westphal : mouvements de vergence
Où se produit la réception de la vision - La rétine
[modifier | modifier le wikicode]Avant d'être transmises, les ondes EM entrantes passent à travers la cornée, le cristallin et la macula. Ces structures agissent également en tant que filtres pour réduire les ondes EM indésirables, protégeant ainsi l’œil de rayonnements nocifs. La réponse de filtrage de chacun de ces éléments est visible dans la figure Filtrage de la lumière effectuée par la cornée, le cristallin et l'épithélium pigmentaire. Comme on peut le constater, la cornée atténue les longueurs d'onde les plus basses, laissant les longueurs d'onde les plus hautes presque intactes. Le cristallin bloque environ 25 % des ondes EM inférieures à 400 nm et plus de 50 % des ondes EM inférieures à 430 nm. Enfin, l'éphithélium pigmentaire, dernière étape de filtrage avant la photoréception, affecte environ 30 % des ondes EM entre 430 et 500 nm.
La partie de l’œil qui marque la transition entre la région non photosensible et la région photosensible, est appelée ora serrata. La région photosensible est appelée la rétine, qui est la structure sensorielle du fond de l’œil. La rétine se compose de plusieurs couches présentées ci-dessous avec des millions de photorécepteurs, appelés bâtonnets et cônes, qui réagissent aux rayons lumineux et les convertissent en impulsions électriques. La transmission de ces impulsions part des cellules ganglionnaires et se fait par le nerf optique, la seule voie par laquelle l'information quitte l’œil.
Une illustration conceptuelle de la structure de la rétine est présentée à droite. Comme on peut le constater, il existe cinq types de cellules principales :
• cellules photoréceptrices
• cellules horizontales
• cellules bipolaires
• cellules d'amécrine
• cellules ganglionnaires
Les cellules photoréceptrices peuvent être subdivisées en deux sous-catégories : les bâtonnets et les cônes. Dans la plupart des régions de la rétine, les cônes sont beaucoup moins nombreux que les bâtonnets mais il en existe une énorme concentration dans la macula, en particulier dans sa partie centrale appelée fovéa. Dans cette région centrale, chaque cône photosensible est connecté à une cellule ganglionnaire. De plus, les cônes de cette région sont légèrement plus petits que les autres ; on observe donc d'autant plus de cônes pour cette zone. C'est grâce à ce ratio et à la forte densité de cônes que nous avons l'acuité visuelle la plus élevée.

Il existe chez l'être humain trois types de cônes, chacun étant sensible à un intervalle spécifique de longueurs d'onde. Les cônes se distinguent par la présence de trois types de pigments appelés photopsines, respectivement sensibles aux longueurs d'onde des lumières rouge, bleue ou verte. Nous avons donc des cônes « bleus », « verts » et « rouges », également appelés cônes S, M et L en raison de leur sensibilité respective aux longueurs d'onde courtes, moyennes et longues. Cela se compose d'une protéine appelée opsine et d'un chromophore lié appelé rétinal. Les principales composantes de la cellule conique sont le terminal synaptique, les segments interne et externe, le noyau interne et les mitochondries.
Les sensibilités spectrales des 3 types de cônes:
• Les cônes S sont sensibles aux ondes courtes, c'est-à-dire à la lumière bleu-violet. La longueur d'onde d'absorption maximale pour les cônes S est de 420 nm.
• Les cônes M sont sensibles à la lumière allant de bleu-vert à jaune. Dans ce cas, la longueur d'onde d'absorption maximale est de 535 nm.
• Les cônes L sont sensibles à la lumière allant de jaune à rouge. La longueur d'onde d'absorption maximale est de 565 nm.
La couche interne contient les organites, le noyau et les organelles de la cellule. Le pigment est situé dans la couche externe, attaché à la membrane sous forme de protéines transmembranaires de la membrane cellulaire qui forment les disques membraneux, clairement visibles sur la figure présentant la structure de base des cellules à cône et à tige. Les disques maximisent la zone de réception des cellules. Les photorécepteurs à cône de nombreux vertébrés contiennent des organites sphériques appelées gouttelettes d'huile, censées constituer des filtres intra-oculaires pouvant servir à augmenter le contraste, à réduire l'éblouissement et à atténuer les aberrations chromatiques causées par le gradient (???), de la taille d'une mitochondrie de la périphérie aux centres.
Les bâtonnets ont une structure similaire à celle des cônes mais ils contiennent à la place un pigment appelé rhodopsine qui leur permet de percevoir la lumière de faible intensité avec une sensibilité 100 fois plus grande que celle des cônes. La rhodopsine est le seul pigment que l'on trouve dans les bâtonnets humains. On la retrouve sur la face externe de l'épithélium pigmentaire, qui, de la même manière que les cônes, maximise la zone d'absorption en utilisant une structure de disque. De la même manière que les cônes, la borne synaptique de la cellule la relie à une cellule bipolaire et les segments interne et externe sont reliés par du cilium.
La rhodopsine absorbe la lumière entre 400 et 600 nm, avec une absorption maximale vers 500 nm. Cette longueur d'onde correspond à la lumière bleu verdâtre, ce qui signifie que la nuit, les couleurs bleues apparaissent plus intenses que les rouges.
Les ondes électromagnétiques dont la longueur d'onde est en dehors de la plage de 400 à 780 nm ne sont détectées ni par des bâtonnets ni par des cônes, et sont donc habituellement invisibles pour les êtres humains.
Remarque : il ne faut toutefois pas oublier que les valeurs de l'efficacité lumineuse, qui caractérise l'intensité de la sensation ressentie par rapport à l'énergie reçue, ne sont que des moyennes considérées pour des raisons pratiques comme une « norme ». En fait, de nombreuses personnes, surtout des hommes, ont une vision des couleurs qui s'écarte de ces moyennes. Par ailleurs, des lumières situées en-dehors des intervalles « normaux », comme la raie ultra-violette du mercure à 365 nm, peuvent être perçues si elles sont suffisamment intenses.
Les cellules horizontales occupent la couche nucléaire interne de la rétine. Il existe deux types de cellules horizontales et les deux types hyper-polarisent en réponse à la lumière, c'est-à-dire qu'ils deviennent plus négatifs (???). Le type A consiste en un sous-type appelé HII-H2 qui interagit principalement avec les cônes en S. Les cellules de type B ont un sous-type appelé HI-H1, qui comporte un arbre à dendrites et un axone. Le premier est en contact principalement avec les cellules à cônes M et L et les dernières cellules à tige (???).
Les contacts avec les cônes sont établis principalement par des synapses prohibitives, tandis que les cellules elles-mêmes sont reliées à un réseau avec des jonctions.
Les cellules bipolaires ont des dendrites uniques dans la couche plexiforme externe et le péricaryon, leurs corps cellulaires se trouvent dans la couche nucléaire interne. Les dendrites s'interconnectent exclusivement avec des cônes et des bâtonnets et nous pouvons distinguer une cellule bipolaire à un bâtonnet ainsi que neuf ou dix cellules bipolaires à cône. Ces cellules se ramifient avec des cellules amacrines ou ganglionnaires dans la couche plexiforme interne en utilisant un axone. Les cellules bipolaires à tige se connectent par des synapses à triades ou à 18 à 70 cellules de tige. Leurs axones s'étendent autour des terminaisons synaptiques de la couche plexiforme interne, qui contiennent des synapses à ruban et entrent en contact avec une paire de processus cellulaires dans les synapses de dyade. Ils sont connectés aux cellules ganglionnaires avec des liaisons de cellules amacrines AII.


Les cellules d'amécrine peuvent être trouvées dans la couche nucléaire interne et dans la couche de cellules ganglionnaires de la rétine, parfois aussi dans la couche plexiforme interne, où elles agissent en tant que modulatrices de signal. Elles ont été classées en champs étroits, petits champs, champs moyens ou champs larges en fonction de leur taille. Cependant, il existe de nombreuses classifications conduisant à plus de 40 types différents de cellules amécrines.
Les cellules ganglionnaires sont les émetteurs terminaux du signal visuel allant de la rétine au cerveau. Les cellules ganglionnaires les plus courantes dans la rétine sont les cellules ganglionnaires de petite taille et les cellules ganglionnaires du parasol. Après avoir traversé toutes les couches rétiniennes, le signal est transmis à ces cellules qui constituent l'étape finale de la chaîne de traitement du signal par la rétine. Toutes les informations sont rassemblées ici, transmises aux fibres nerveuses rétiniennes et aux nerfs optiques. Le point où les axones ganglionnaires fusionnent pour créer un nerf optique s'appelle le disque optique. Ce nerf est construit principalement à partir des axones ganglionnaires de la rétine. La majorité des axones transmettent des données au noyau géniculé latéral, qui est un lien de terminaison pour la plupart des parties du nerf et qui transmet l'information au cortex visuel. Certaines cellules ganglionnaires réagissent également à la lumière mais comme cette réponse est plus lente que celle des bâtonnets et des cônes, on pense qu'elle est liée à la détection des niveaux de lumière ambiante et au réglage de l'horloge biologique.
Le traitement du signal
[modifier | modifier le wikicode]Comme mentionné précédemment, la rétine est le composant principal de l’œil car elle contient toutes les cellules sensibles à la lumière. Sans elle, l’œil serait comparable à un appareil photo dépourvu de film ou de capteur. Cette partie montre comment la rétine perçoit la lumière, comment le signal optique est transmis au cerveau puis comment le cerveau traite le signal pour élaborer suffisamment d'informations en vue de la prise de décision.
La création initiale du signal - Les cellules photosensibles
[modifier | modifier le wikicode]La vision commence invariablement par la lumière frappant les cellules photosensibles présentes dans la rétine. Les pigments visuels absorbant la lumière, une variété d'enzymes et de transmetteurs dans des bâtonnets et des cônes rétiniens vont initier la conversion des stimuli électromagnétiques visibles en impulsions électriques, selon un processus appelé transduction photoélectrique. En prenant des bâtonnets à titre d'exemple, l'EM visible visible frappe les molécules de rhodopsine, molécules transmembranaires présentes dans la structure du disque externe des bâtonnets. Chaque molécule de rhodopsine consiste en un groupe d'hélices appelé opsine, qui enveloppent et entourent le rétinal 11-cis, qui est la partie de la molécule qui changera en raison de l'énergie des photons entrants. Dans les molécules biologiques, les fractions ou parties de molécules qui provoqueront des changements de conformation dus à cette énergie sont parfois appelées chromophores. Le 11-cis rétinien se redresse en réponse à l'énergie entrante, se transformant en rétinien (rétinien tout trans), ce qui force les hélices à opsine à se séparer, provoquant la découverte de sites réactifs particuliers. Cette molécule de rhodopsine "activée" est parfois appelée métarhodopsine II. À partir de ce moment, même si la stimulation par la lumière visible cesse, la réaction se poursuivra. La métarhodopsine II peut alors réagir avec environ 100 molécules d'une protéine Gs appelée transduction, ce qui aboutit alors à l'as and ßs. Ensuite le GDP est converti en GTP. Le GTP as activé se lie ensuite à la cGMP-phosphodiestérase (PDE), supprimant les fonctions normales d'échange d'ions, ce qui entraîne une faible concentration en cytosol des ions cationiques, et par conséquent une modification de la polarisation de la cellule.
La réaction de transduction photoélectrique naturelle possède un incroyable pouvoir d'amplification. Une seule molécule de rhodopsine rétinienne activée par un seul quantum de lumière provoque l'hydrolyse d'un maximum de 106 molécules de GMPc par seconde.
La phototransduction
[modifier | modifier le wikicode]1.Un photon lumineux interagit avec le rétinal dans un photorécepteur. Le rétinal subit une isomérisation, passant de la configuration 11-cis à la configuration tout-trans.
2.Le rétinal ne s'adapte plus dans le site de liaison à l'opsine.
3.L'opsin subit donc un changement de conformation en métarhodopsine II.
4.La métarhodopsine II est instable et se scinde, donnant de l'opsine et du rétinal tout-trans.
5.L'opsine active la transducine régulatrice de la protéine. Ce qui provoque la dissociation de la transducine se dissocie de son PIB lié et lie le GTP, puis la sous-unité alpha de la transducine se dissocie des sous-unités bêta et gamma, le GTP étant toujours lié à la sous-unité alpha.
6.Le complexe sous-unité alpha-GTP active la phosphodiesterase.
7.La phosphodiesterase décompose le GMPc en 5'-GMP. Cela diminue la concentration de GMPc et donc la fermeture des canaux sodiques.
8.La fermeture des canaux sodiques provoque une hyperpolarisation de la cellule en raison du courant de potassium en cours.
9.L'hyperpolarisation de la cellule provoque la fermeture des canaux calciques dépendants du voltage.
10.Lorsque le niveau de calcium dans la cellule photoréceptrice diminue, la quantité de glutamate de neurotransmetteur libérée par la cellule diminue également. Cela est dû au fait que le calcium est nécessaire pour que les vésicules contenant du glutamate fusionnent avec la membrane cellulaire et libèrent leur contenu.
11.Une diminution de la quantité de glutamate libérée par les photorécepteurs provoque une dépolarisation des cellules bipolaires centrées (cellules bipolaires à bâtonnets et à cônes) et une hyperpolarisation des cellules bipolaires à cône éteint.
Sans stimulation électromagnétique visible, les cellules en bâtonnets contenant un cocktail d'ions, de protéines et d'autres molécules présentent des différences de potentiel membranaire d'environ -40 mV. Comparé aux autres cellules nerveuses, ce potentiel est assez élevé (-65 mV). Dans cet état, le glutamate de neurotransmetteur est libéré en continu des terminaisons axonales et absorbé par les cellules bipolaires voisines. Avec l'EM visible et la réaction en cascade mentionnée précédemment, la différence de potentiel chute à -70 mV. Cette hyper-polarisation de la cellule provoque une réduction de la quantité de glutamate libéré, affectant ainsi l'activité des cellules bipolaires et, par la suite, les étapes suivantes de la voie optique.
Des processus similaires existent dans les cellules du cône et dans les cellules ganglionnaires photosensibles, mais utilisent des opsines différentes. Les photopsines I à III (respectivement vertes-jaunâtres, vertes et bleues-violettes) se trouvent dans les trois différentes cellules du cône et on peut trouver la mélanopsine (bleue) dans les cellules ganglionnaires photosensibles.

Le traitement du signal dans la rétine
[modifier | modifier le wikicode]Différentes cellules bipolaires réagissent différemment aux modifications du glutamate libéré. Les cellules bipolaires ON et OFF sont utilisées pour former le flux de signal direct des cônes aux cellules bipolaires. Les cellules bipolaires ON vont se dépolariser par stimulation électromagnétique visible et les cellules ganglionnaires ON correspondantes seront activées. D'autre part, les cellules bipolaires OFF sont hyper polarisées par la stimulation EM visible et les cellules ganglionnaires OFF sont inhibées. C'est la voie de base du flux de signal direct. Le flux de signal latéral commencera à partir des bâtonnets, puis ira aux cellules bipolaires, puis aux cellules amacrines, et les cellules bipolaires OFF inhibées par les cellules Rod-amacrine et les cellules bipolaires ON seront stimulées via une synapse électrique, après toutes les étapes prévues, le signal arrivera aux cellules ganglionnaires activées ou désactivées et ainsi l'ensemble du trajet du flux de signal latéral est établi.

Lorsque le potentiel d'action (PA) est activé, les cellules ganglionnaires seront déclenchées par le stimulus EM visible. La fréquence AP augmentera lorsque le potentiel du capteur augmente. En d'autres termes, AP dépend de l'amplitude du potentiel du capteur. La région des cellules ganglionnaires où les effets stimulants et inhibiteurs influencent la fréquence des AP s'appelle le champ récepteur (RF). Autour des cellules ganglionnaires, le RF est généralement composé de deux régions: la zone centrale et la zone périphérique en forme d'anneau. Ils se distinguent lors de l'adaptation visible aux EM. Une stimulation électromagnétique visible sur la zone centrale pourrait entraîner une augmentation de la fréquence du PA et la stimulation sur la zone périphérique diminuerait la fréquence du PA. Lorsque la source de lumière est éteinte, l'excitation se produit. Le nom du champ ON (champ central ON) fait donc référence à ce type de région. Bien entendu, la RF des cellules ganglionnaires OFF agit dans le sens opposé et est donc appelée "champ OFF" (champ central OFF). Les RF sont organisés par les cellules horizontales. L'impulsion sur la région périphérique sera impulsée et transmise à la région centrale, là où le contraste appelé stimulus est formé. Cette fonction rendra l'obscurité plus sombre et la lumière plus brillante. Si tout le RF est exposé à la lumière. l'impulsion de la région centrale prédominera.
La transmission du signal au cortex
[modifier | modifier le wikicode]Comme mentionné précédemment, les axones des cellules ganglionnaires convergent vers le disque optique de la rétine, formant le nerf optique. Ces fibres sont positionnées à l'intérieur du faisceau dans un ordre spécifique. Les fibres de la zone maculaire de la rétine se trouvent dans la partie centrale et celles de la moitié temporale de la rétine occupent la partie périphérique. Une décussation ou un croisement partiel se produit lorsque ces fibres se trouvent en dehors de la cavité oculaire. Les fibres des moitiés nasales de chaque rétine se croisent vers les moitiés opposées et s'étendent jusqu'au cerveau.
Celles des moitiés temporales restent à la périphérie. Ce croisement partiel s'appelle le chiasma optique, et les nerfs optiques au-delà de ce point s'appellent les voies optiques, principalement pour les distinguer des nerfs rétiniens simples. Le croisement partiel a pour fonction de transmettre le champ visuel de la main droite produit par les deux yeux à la moitié gauche du cerveau uniquement et inversement. Par conséquent, les informations de la moitié droite du corps et du champ visuel droit sont toutes transmises à la partie gauche du cerveau lorsqu'elles atteignent la partie postérieure de l'avant-cerveau (diencephale).

Le relais d'information entre les fibres du tractus optique et les cellules nerveuses se produit dans les corps géniculés latéraux, la partie centrale du traitement du signal visuel, situé dans le thalamus du cerveau. C'est d'ici que l'information est transmise aux cellules nerveuses vers le cortex occipital du côté correspondant du cerveau. Les connexions de la rétine au cerveau peuvent être séparées en une "voie parvocellulaire" et une "voie magnocellulaire". Les voies parvocellulaires signalent la couleur et les détails fins, tandis que les voies magnocellulaires détectent les stimuli en mouvement rapide.
Les signaux des appareils photo numériques standard correspondent approximativement à ceux de la voie parvocellulaire. Pour simuler les réponses des voies parvocellulaires, les chercheurs ont développé des systèmes sensoriels neuromorphes, qui tentent d'imiter le calcul basé sur les pointes dans les systèmes neuronaux. Pour ce faire ils utilisent un schéma appelé "représentation d'adresse-événement" pour la transmission du signal dans les systèmes électroniques neuromorphes (Liu et Delbruck 2010 [1]).
Sur le plan anatomique, les cellules ganglionnaires de Magno et de Parvo rétiniennes projettent respectivement 2 couches magnocellulaires ventrales et 4 couches parvocellulaires dorsales du noyau géniculé latéral (LGN). Chacune des six couches de LGN reçoit des entrées de l’œil ipsilatéral ou contralatéral, c'est-à-dire que les cellules ganglionnaires de l’œil gauche se croisent et se projettent sur les couches 1, 4 et 6 de la LGN droite. Et les cellules ganglionnaires de l’œil droit se projettent (sans se croiser) sur les couches 2,3 et 5. À partir de là, les informations provenant de l’œil droit et de l’œil gauche sont séparées.
Bien que la vision humaine soit combinée par deux moitiés de la rétine et que le signal soit traité par les hémisphères cérébraux opposés, le champ visuel est considéré comme une unité lisse et complète. Par conséquent, les deux aires corticales visuelles sont considérées comme étant intimement liées. Cette connexion, appelée corpus callosum, est constituée de neurones, d'axones et de dendrites. Comme les dendrites établissent des connexions synaptiques avec les points correspondants des hémisphères, la simulation électrique de tout point d'un hémisphère indique la simulation du point interconnecté de l'autre hémisphère. La seule exception à cette règle est le cortex visuel primaire.
Les synapses sont fabriquées par le tractus optique dans les couches respectives du corps géniculé latéral. Ensuite, ces axones de ces cellules nerveuses de troisième ordre sont transmis à la fissure calcarine dans chaque lobe occipital du cortex cérébral. Comme les bandes des fibres blanches et des paires d'axones des cellules nerveuses de la rétine le traversent, on l'appelle le cortex strié, qui est d'ailleurs notre principal cortex visuel, parfois appelé V1. À ce stade, les impulsions des yeux séparés convergent vers les neurones corticaux communs, ce qui permet ensuite l'entrée complète des deux yeux dans une région afin d'être utilisée pour la perception et la compréhension. La reconnaissance des formes est une fonction très importante de cette partie du cerveau, les lésions causant des problèmes de reconnaissance visuelle ou de vision aveugle.
Si l'on se fonde sur la manière ordonnée selon laquelle les fibres du tractus optique transmettent des informations aux corps géniculés latéraux et ensuite vers la zone striée, et si un seul point de stimulation sur la rétine était trouvé, la réponse qui s'est produite électriquement à la fois dans le corps géniculé latéral et le cortex strié se trouverait dans une petite région de la tache rétinienne. Ceci est la manière évidente de traiter le signal point à point. Et si toute la rétine est entièrement stimulée, les réponses se produiront à la fois sur les deux corps géniculés latéraux et sur la zone de matière grise du cortex strié. Il est possible de dresser une carte de cette région du cerveau jusqu'aux champs rétiniens, ou plus généralement jusqu'aux champs visuels.
Toute autre étape dans cette voie sort du cadre de ce livre. Soyez assuré que de nombreux autres niveaux et centres existent, en vous concentrant sur des tâches spécifiques, telles que la couleur, les orientations, les fréquences spatiales, les émotions, etc.
Le traitement de l'information dans le système visuel
[modifier | modifier le wikicode]Équipé d'une compréhension plus fine de certains des concepts les plus importants du traitement du signal dans le système visuel, la compréhension ou la perception de l'information sensorielle traitée est la dernière pièce importante du puzzle. La perception visuelle est le processus de traduction des informations reçues par les yeux dans la compréhension de l'état extérieur des choses. Cela nous rend conscients du monde qui nous entoure et nous permet de mieux le comprendre. Sur la base de la perception visuelle, nous apprenons des modèles que nous appliquons plus tard dans la vie et nous prenons des décisions en fonction de ces informations perçues. En d'autres termes, notre survie dépend de la perception. Le champ de la perception visuelle a été divisé en différents sous-champs, en raison du fait que le traitement est trop complexe et nécessite divers mécanismes spécialisés pour percevoir ce qui est vu. Ces sous-champs incluent: la perception des couleurs, la perception du mouvement, la perception de la profondeur, la reconnaissance des visages, etc.
Le cortex visuel des primates
[modifier | modifier le wikicode]Malgré la puissance de calcul sans cesse croissante des systèmes électroniques, il existe encore de nombreuses tâches pour lesquelles les animaux et les humains dépassent de loin les ordinateurs, notamment la perception et la contextualisation de l'information. L'ordinateur classique, que ce soit celui de votre téléphone ou celui d'un supercalculateur occupant toute la pièce, est essentiellement un calculateur de chiffres. Il peut effectuer une quantité incroyable de calculs en un temps infime. Ce qui lui manque, c'est de créer des abstractions de l'information avec laquelle il travaille. Si vous connectez une caméra à votre ordinateur, l'image qu'elle perçoit n'est qu'une grille de pixels, un tableau de nombres en 2 dimensions. Un humain reconnaîtrait immédiatement la géométrie de la scène, les objets de la photo et peut-être même le contexte de ce qui se passe. Cette capacité que nous avons, nous est fournie par une machinerie biologique dédiée - le système visuel du cerveau.
Il traite tout ce que nous voyons de manière hiérarchique, depuis les fonctionnalités les plus simples de l'image jusqu'aux plus complexes, en passant par la classification des objets en catégories. On dit donc que le système visuel a une hiérarchie profonde. La hiérarchie profonde du système visuel des primates a incité les informaticiens à créer des modèles de réseaux de neurones artificiels comportant également plusieurs couches, chacune d'elles créant des généralisations plus élevées des données d'entrée.
Environ la moitié du néocortex humain est dédiée à la vision. Le traitement des informations visuelles s'effectue sur au moins 10 niveaux fonctionnels. Les neurones dans les premières zones visuelles extraient des caractéristiques d'image simples sur de petites régions locales de l'espace visuel. Lorsque les informations sont transmises aux zones visuelles supérieures, les neurones répondent à des caractéristiques de plus en plus complexes. Avec des niveaux plus élevés de traitement de l'information, les représentations deviennent plus invariantes - moins sensibles à la taille, à la rotation ou à la position exactes des caractéristiques. De plus, la taille du champ récepteur des neurones dans les zones visuelles supérieures augmente, indiquant qu'ils sont réglés sur des caractéristiques d'image plus globales. Cette structure hiérarchique permet un calcul efficace: différentes zones visuelles supérieures peuvent utiliser les mêmes informations calculées dans les zones inférieures. La description générique de la scène qui est faite dans les premières zones visuelles est utilisée par d'autres parties du cerveau pour effectuer diverses tâches, telles que la reconnaissance et la catégorisation d'objets, la saisie, la manipulation, la planification de mouvements, etc.
La vision sous-corticale
[modifier | modifier le wikicode]
Le traitement neuronal de l'information visuelle commence déjà avant toute structure corticale. Les photorécepteurs sur la rétine détectent la lumière et envoient des signaux aux cellules ganglionnaires de la rétine. La taille du champ récepteur d'un photorécepteur est d'un centième de degré (un champ récepteur d'environ un degré correspond à peu près à la taille de votre pouce lorsque vous avez le bras tendu devant vous). Le nombre d'entrées dans une cellule ganglionnaire et donc la taille de son champ récepteur dépendent de son emplacement: au centre de la rétine, elle reçoit des signaux provenant de cinq récepteurs seulement, tandis qu'à la périphérie, une seule cellule peut contenir plusieurs milliers d'entrées. Cela implique que la résolution spatiale la plus élevée se situe au centre de la rétine, également appelée fovéa. En raison de cette propriété, les primates possèdent un mécanisme de contrôle du regard qui dirige la vue de sorte que les caractéristiques intéressantes se projettent sur la fovéa.
Les cellules ganglionnaires sont réglées de manière sélective pour détecter diverses caractéristiques de l'image, telles que le contraste de luminance, le contraste de couleur, la direction et la vitesse de déplacement. Toutes ces caractéristiques constituent les informations principales utilisées ultérieurement dans le pipeline de traitement. S'il existe des stimuli visuels qui ne sont pas détectables par les cellules ganglionnaires, ils ne sont également disponibles pour aucun espace visuel cortical.
Les cellules ganglionnaires se projettent dans une région du thalamus appelée noyau géniculé latéral (LGN), qui à son tour relaie les signaux au cortex. Il n'y a pas de calcul significatif connu dans LGN - il y a presque une correspondance un-à-un entre le ganglion rétinien et les cellules LGN. Cependant, seulement 5% des entrées de LGN proviennent de la rétine - toutes les autres entrées sont des projections à rétroaction corticale. Bien que le système visuel soit souvent considéré comme un système à feed-forward, les connexions de rétroaction récurrentes ainsi que les connexions latérales sont une caractéristique commune observée dans tout le cortex visuel. Le rôle de la rétroaction n'est pas encore complètement compris, mais il est proposé de l'attribuer à des processus tels que l'attention, les attentes, l'imagination et le remplissage des informations manquantes.
La vision corticale
[modifier | modifier le wikicode]Le cortex visuel peut être divisé en trois grandes parties: la partie occipitale qui reçoit les entrées de LGN puis envoie les sorties par le flux dorsal et ventral. La partie occipitale comprend les zones V1-V4 et MT, qui traitent divers aspects de l'information visuelle et donne lieu à la représentation générique des scènes. La voie dorsale est impliquée dans l'analyse de l'espace et dans la planification de l'action. La voie ventrale participe à la reconnaissance et à la catégorisation des objets.
V1 est la première zone corticale qui traite les informations visuelles. Elle est sensible aux contours extérieurs, aux réseaux, aux fins de ligne, au mouvement, à la couleur et aux disparités (différence angulaire entre les projections d'un point sur les rétines gauche et droite). L'exemple le plus simple du traitement hiérarchique ascendant est la combinaison linéaire des entrées de plusieurs cellules ganglionnaires avec des champs récepteurs centraux pour créer la représentation d'une barre. Ceci est fait par les cellules simples de V1 et a été décrit pour la première fois par les neuroscientifiques renommés Hubel et Wiesel. Ce type d'intégration d'informations implique que les cellules simples sont sensibles à l'emplacement exact de la barre et ont un champ de réception relativement petit. Les cellules complexes de V1 reçoivent les entrées des cellules simples et, bien que répondant également à des modèles orientés de manière linéaire, elles ne sont pas sensibles à la position exacte de la barre et ont un champ de réception plus grand. Le calcul présenté à cette étape pourrait être une opération de type MAX produisant des réponses d'amplitude similaire à la plus grande des réponses des stimuli. Certaines cellules simples et complexes peuvent également détecter la fin d'une barre et une fraction des cellules V1 sont également sensibles au mouvement local dans leurs champs récepteurs respectifs.
La zone V2 offre une représentation plus sophistiquée des contours, notamment des contours définis par une texture, des contours illusoires et des contours qui possèdent une bordure. V2 s'appuie également sur la détection de la disparité absolue dans V1 et présente des cellules sensibles à la disparité relative, qui correspond à la différence entre les disparités absolues de deux points de l'espace. La zone V4 reçoit les entrées de V2 et la zone V3, mais on sait très peu de choses sur le calcul effectué dans V3. La zone V4 contient des neurones sensibles aux contours de courbure différente et aux sommets d'angles particuliers. Une autre caractéristique importante est le codage de la teinte invariante de la luminance. Cela contraste avec V1 où les neurones réagissent à une opposition de couleur le long des deux axes principaux (rouge-vert et jaune-bleu) plutôt que la couleur réelle. La V4 est ensuite transmise au flux ventral vers le cortex temporal inférieur (IT), ce qui a été démontré par des études sur les lésions comme étant essentiel pour la discrimination d'objet.
Le cortex temporal inférieur: différenciation des objets
[modifier | modifier le wikicode]
Le cortex temporal inférieur (IT) est divisé en deux zones: TEO et TE. La région TEO intègre des informations sur les formes et les positions relatives d'éléments aux multiples contours et comprend principalement des cellules répondant à de simples combinaisons de caractéristiques. La taille du champ récepteur des neurones TEO est d'environ 3 à 5 degrés. Area TE comporte des cellules avec des champs récepteurs beaucoup plus grands (10 à 20 degrés) qui correspondent aux visages, aux mains et aux configurations complexes. Les cellules de TE répondent à des caractéristiques visuelles qui sont une généralisation plus simple de l'objet d'intérêt, mais plus complexe que de simples barres ou points. Ceci a été démontré en utilisant une méthode de réduction de stimulus par Tanaka et al. dans laquelle une réponse à un objet est tout d'abord mesurée, puis l'objet est remplacé par des représentations plus simples jusqu'à ce que la caractéristique critique à laquelle répondent les neurones TE soit réduite.
Il semble que les neurones informatiques combinent diverses caractéristiques de complexité moyenne à partir des niveaux inférieurs du flux ventral pour créer des modèles de parties d'objet. Les neurones de TE qui sont sélectifs pour des objets spécifiques doivent remplir deux exigences apparemment contradictoires: la sélectivité et l'invariance. Ils doivent faire la distinction entre différents objets au moyen de la sensibilité aux caractéristiques des images rétiniennes. Cependant, le même objet peut être vu sous différents angles et à différentes distances dans différentes conditions d'éclairage, ce qui donne des images rétiniennes extrêmement différentes du même objet.
Si l'on considère toutes ces images comme équivalentes, il faut dériver des caractéristiques invariantes qui résistent à certaines transformations, telles que des changements de position, d'éclairage, de taille sur la rétine, etc. Les neurones dans la zone TE montrent une invariance de position et de taille ainsi que dans l'occlusion partielle, de la position en profondeur et direction de l'éclairage. Il a été démontré que la rotation en profondeur avait la plus faible invariance, à l'exception si l'objet est un visage humain.
Les catégories d'objets ne sont pas encore explicitement présentes dans la zone TE. Un neurone peut généralement correspondre à plusieurs exemples de la même catégorie (par exemple, des images d'arbres), mais pas à tous.
Il peut également correspondre à des exemples de différentes catégories (par exemple, les arbres et les non-arbres). La reconnaissance et la classification des objets impliquent très probablement un échantillonnage d'une population plus large de neurones TE ainsi que la réception d'entrées d'autres zones du cerveau, par exemple celles responsables de la compréhension du contexte de la scène. Des expériences de lecture récentes ont montré que des classificateurs statistiques (par exemple, des machines à vecteurs de support) peuvent être formés à la classification d'objets en fonction des réponses d'un petit nombre de neurones TE. Par conséquent, une population composée essentiellement de neurones TE peut en principe indiquer de manière fiable grâce à la combinaison de leur activité. De manière intéressante, il existe également des rapports sur des neurones hautement sélectifs dans le lobe temporal médial qui répondent à des signaux très spécifiques, par exemple à la tour de Pise, dans différentes photos ou au visage d'une personne donnée.
L'apprentissage dans le système visuel
[modifier | modifier le wikicode]L'apprentissage peut altérer la sélectivité visuelle des neurones, ce qui a pour effet de rendre plus difficile l'apprentissage aux niveaux hiérarchiques supérieurs. Il n'existe aucune preuve d'apprentissage dans la rétine et les schémas d'orientation de V1 semblent être génétiquement largement prédéterminées. Cependant, la pratique de l'identification d'orientation améliore le codage d'orientation dans les neurones V1 en augmentant la pente de la courbe de réglage. Des effets similaires mais plus importants ont été observés dans V4. En zone TE, relativement peu d'entraînement visuel a produit des effets physiologiques notables sur la perception visuelle, aussi bien au niveau d'une cellule unique, qu'en IRMf. Par exemple, transformer deux objets l'un en l'autre augmente leur similarité perçue. Globalement, il semble que même le cortex visuel adulte soit considérablement plastique et le niveau de plasticité peut être considérablement accru, par exemple en administrant des médicaments spécifiques ou en vivant dans un environnement enrichi.
Les réseaux d'apprentissage profond
[modifier | modifier le wikicode]À l'instar de la hiérarchie profonde du système visuel des primates, les architectures d'apprentissage approfondi tentent de modéliser des abstractions de haut niveau des données d'entrée en utilisant plusieurs niveaux de transformations non linéaires. Le modèle proposé par Hubel et Wiesel dans lequel les informations sont intégrées et propagées en cascade de la rétine et du LGN aux cellules simples et complexes en V1 a inspiré la création de l'une des premières architectures d'apprentissage en profondeur, le néocognitron -un modèle de réseau neuronal artificiel multicouche. Il a été utilisé pour différentes tâches de reconnaissance de formes, y compris la reconnaissance de caractères manuscrits. Cependant, la formation du réseau a pris beaucoup de temps (de l'ordre de plusieurs jours) et, depuis sa création dans les années 1980, l'apprentissage en profondeur n'a pas attiré l'attention jusqu'au milieu des années 2000 avec l'abondance des données numériques et l'invention des algorithmes d'entraînement accéléré. Les réseaux de neurones profonds se sont révélés très efficaces dans des tâches qui, il n'y a pas si longtemps, semblaient être réservées aux humains, telles que la reconnaissance des visages de personnes particulières sur des photos, la compréhension du discours humain (dans une certaine mesure) et la traduction de textes à partir de langues étrangères . En outre, ils se sont révélés très utiles dans l'industrie et la science pour rechercher des médicaments potentiels, cartographier les réseaux de neurones réels dans le cerveau et prédire les fonctions des protéines. Il faut noter que l'apprentissage en profondeur n'est inspiré que très vaguement par le cerveau et constitue bien davantage une réussite du domaine de l'informatique / apprentissage automatique que de celui des neurosciences. Les parallèles de base sont que les réseaux de neurones profonds sont composés d'unités qui intègrent les entrées d'informations de manière non linéaire (neurones) et s'envoient des signaux (synapses) et qu'il existe différents niveaux de représentations de plus en plus abstraites des données.
Les algorithmes d'apprentissage et les descriptions mathématiques des neurones utilisés dans l'apprentissage en profondeur sont très différents des processus réels se déroulant dans le cerveau. Par conséquent, la recherche en apprentissage en profondeur, tout en donnant une impulsion considérable à une intelligence artificielle plus sophistiquée, ne peut donner que des informations limitées sur le cerveau.
Sources
[modifier | modifier le wikicode]- Apprentissage profond dans le système visuel
- N. Kruger, « Deep Hierarchies in the Primate Visual Cortex: What Can We Learn for Computer Vision? », dans IEEE Transactions on Pattern Analysis and Machine Intelligence, vol. 35, no 8, August 2013, p. 1847–1871 [lien DOI]
- Tomaso Poggio, « (1 novembre 1999) », dans Nature Neuroscience, vol. 2, no 11, 1 November 1999, p. 1019–1025 [lien DOI](1 novembre 1999)
- Expérience d'inhibition des stimuli
- Keiji Tanaka, « Inferotemporal Cortex and Object Vision », dans Annual Review of Neuroscience, vol. 19, no 1, March 1996, p. 109–139 [lien DOI]
- Preuves de l'apprentissage dans le système visuel
- Nuo Li, « Unsupervised Natural Visual Experience Rapidly Reshapes Size-Invariant Object Representation in Inferior Temporal Cortex », dans Neuron, vol. 67, no 6, 23 September 2010, p. 1062–1075 [lien DOI]
- S. Raiguel, « Learning to See the Difference Specifically Alters the Most Informative V4 Neurons », dans Journal of Neuroscience, vol. 26, no 24, 14 June 2006, p. 6589–6602 [lien DOI]
- A Schoups, « Practising orientation identification improves orientation coding in V1 neurons. », dans Nature, vol. 412, no 6846, 2 August 2001, p. 549-53 [lien PMID]
- Vue d'ensemble récente et accessible de l'apprentissage profond
- Nicola Jones, « Computer science: The learning machines », dans Nature, vol. 505, no 7482, 8 January 2014, p. 146–148 [lien DOI]
Perception du mouvement
[modifier | modifier le wikicode]La perception du mouvement est le processus de déduction de la vitesse et de la direction des objets en mouvement. La zone V5 chez l'homme et la zone MT (région temporale moyenne) chez les primates sont responsables de la perception corticale du mouvement. La zone V5 fait partie du cortex extrastrié, qui est la région située dans la région occipitale du cerveau, à côté du cortex visuel principal. La zone V5 a pour fonction de détecter la vitesse et la direction des stimuli visuels et d’intégrer les signaux de mouvement visuels locaux au mouvement global. La région V1 ou cortex visuel primaire est située dans le lobe occipital du cerveau dans les deux hémisphères. Il traite la première étape du traitement cortical de l'information visuelle. Cette zone contient une carte complète du champ visuel couvert par les yeux. La différence entre la zone V5 et la zone V1 (Cortex visuel principal) est que la zone V5 peut intégrer le mouvement de signaux locaux ou de parties individuelles d'un objet dans le mouvement global d'un objet entier. La zone V1, en revanche, répond au mouvement local qui se produit dans le champ récepteur. Les estimations de ces nombreux neurones sont intégrées dans la zone V5.
Le mouvement est défini comme des changements dans l'illumination de la rétine dans l'espace et dans le temps. Les signaux de mouvement sont classés en mouvements de premier ordre et en mouvements de second ordre. Ces types de mouvement sont décrits brièvement dans les paragraphes suivants.

La perception du mouvement de premier ordre fait référence au mouvement perçu lorsque deux stimuli visuels ou plus s’allument et s’éteignent avec le temps et produisent des perceptions de mouvement différentes. Le mouvement de premier ordre est également appelé mouvement apparent, et est utilisé à la télévision et au cinéma. Le mouvement Beta en est un exemple, une illusion dans laquelle les images fixes semblent bouger, même si elles sont en réalité immobiles. Ces images donnent l’apparence d’un mouvement, car elles changent et se déplacent plus rapidement que ce que l’œil peut détecter. Cette illusion d'optique se produit parce que le nerf optique humain réagit à des changements de lumière à dix cycles par seconde, signifiant que tout changement plus rapide que cette fréquence sera enregistré comme un mouvement continu, et non comme des images séparées.
Le mouvement de second ordre fait référence au mouvement qui se produit lorsqu'un contour en mouvement est défini par un contraste, une texture, un scintillement ou toute autre qualité qui n'entraîne pas une augmentation de la luminance ou de l'énergie de mouvement de l'image. Différentes preuves suggèrent que le traitement initial des requêtes de premier ordre et des requêtes de second ordre est effectué par des voies distinctes. Les mécanismes de second ordre ont une résolution temporelle plus faible et se comportent en tant que filtre passe-bas en termes de gamme de fréquences spatiales auxquelles ils répondent. Le mouvement de second ordre produit un effet postérieur de mouvement plus faible. Les signaux de premier et de second ordre sont combinés au sein de la zone V5.
Dans ce chapitre, nous allons analyser les concepts de la perception du mouvement et de l’analyse du mouvement, et expliquer pourquoi ces termes ne doivent pas être utilisés de manière interchangeable. Nous analyserons les mécanismes par lesquels le mouvement est perçu, tels que les capteurs de mouvement et le suivi de charactéristiques. Il existe trois principaux modèles théoriques qui tentent de décrire la fonction des capteurs neuronaux de mouvement. Des tests expérimentaux ont été effectués pour confirmer l'exactitude de ces modèles. Malheureusement, les résultats de ces tests ne sont pas concluants et on peut dire qu'aucun de ces modèles ne décrit complètement le fonctionnement des capteurs de mouvement. Cependant, chacun de ces modèles simule certaines fonctionnalités des capteurs de mouvement. Certaines propriétés de ces capteurs sont décrites. Enfin, ce chapitre présente des illusions de mouvement, qui démontrent que notre sens du mouvement peut être induit en erreur par des facteurs externes statiques qui stimulent les capteurs de mouvement de la même manière que le ferait un vrai mouvement.
Analyse et perception du mouvement
[modifier | modifier le wikicode]Les concepts d'analyse de mouvement et de perception du mouvement sont souvent confondus. La perception et l'analyse du mouvement sont importantes l'une pour l'autre, mais elles ne sont pas identiques.
L'analyse de mouvement fait référence aux mécanismes dans lesquels les signaux de mouvement sont traités. De la même manière que la perception du mouvement ne dépend pas nécessairement des signaux générés par le mouvement des images dans la rétine, l'analyse du mouvement peut ou non conduire à la perception du mouvement. Un exemple de ce phénomène est la vection, qui se produit lorsqu'une personne perçoit qu'elle bouge alors qu'elle est immobile, mais qu’en vérité, l'objet qu'elle observe est en mouvement. La vection montre que le mouvement d'un objet peut être analysé, même s'il n'est pas perçu comme un mouvement provenant de l'objet. Cette définition de l'analyse de mouvement suggère que le mouvement est une propriété fondamentale de l'image. Dans le champ visuel, il est analysé à chaque point. Les résultats de cette analyse sont utilisés pour dériver des informations perceptuelles.
La perception du mouvement désigne le processus d’acquisition de connaissances perceptuelles sur le mouvement d’objets et de surfaces dans une image. Le mouvement est perçu soit par des capteurs locaux délicats dans la rétine, soit par suivi de caractéristiques. Les capteurs de mouvement locaux sont des neurones spécialisés sensibles au mouvement et analogues aux capteurs spécialisés de couleur. Le suivi de charactéristiques est un moyen indirect de percevoir le mouvement. Il consiste à déduire le mouvement des modifications de la position rétinienne des objets au fil du temps. On parle également d'analyse de mouvement de troisième ordre. Le suivi de charactéristiques fonctionne en focalisant l'attention sur un objet particulier et en observant l'évolution de sa position au fil du temps.
Capteurs de mouvement
[modifier | modifier le wikicode]La détection de mouvement est la première étape du traitement visuel. Elle se produit grâce à des processus neuronaux spécialisés, qui répondent aux informations concernant les changements locaux d'intensité des images au fil du temps. Le mouvement est détecté indépendamment des autres propriétés de l'image à chaque point de l'image. Il a été prouvé que des capteurs de mouvement existent et fonctionnent localement à tous les points de l’image. Les capteurs de mouvement sont des capteurs neuronaux dédiés situés dans la rétine et capables de détecter un mouvement produit par deux brefs et faibles flashs lumineux si proches qu'ils ne pourraient pas être détectés par le suivi de caractéristiques. Il existe trois modèles principaux qui tentent de décrire le fonctionnement de ces capteurs spécialisés. Ces modèles sont indépendants les uns des autres et tentent de modéliser les caractéristiques spécifiques de la perception du mouvement. Bien qu’il n’existe pas suffisamment de preuves permettant de penser qu’un de ces modèles représente la façon dont le système visuel (en particulier les capteurs de mouvement) perçoit le mouvement, ils modélisent néanmoins correctement certaines fonctions de ces capteurs.

Le Détecteur Reichardt
Le détecteur Reichardt est utilisé pour modéliser la façon dont les capteurs de mouvement répondent aux signaux de mouvement du premier ordre. Lorsqu'un objet se déplace d'un point A du champ visuel à un point B, deux signaux sont générés: un avant le début du mouvement et un autre après la fin du mouvement. Ce modèle perçoit ce mouvement en détectant les changements de luminance en un point de la rétine et en le corrélant avec un changement de luminance en un autre point à proximité après un court délai. Le détecteur Reichardt fonctionne selon le principe de corrélation (relation statistique qui implique une dépendance). Il interprète un signal de mouvement par corrélation spatio-temporelle des signaux de luminance aux points voisins. Il utilise le fait que deux champs récepteurs situés à différents points de la trajectoire d'un objet en mouvement reçoivent une version décalée du même signal - un diagramme de luminance se déplace le long d'un axe et le signal en un point de l'axe est une version décalée du temps un signal précédent dans l'axe. Le modèle de détecteur Reichardt comporte deux détecteurs voisins séparés dans l'espace. Les signaux de sortie des détecteurs sont multipliés (corrélés) de la manière suivante: un signal multiplié par un deuxième signal qui est la version décalée dans le temps de l'original. La même procédure est répétée mais dans le sens inverse du mouvement (le signal qui a été décalé dans le temps devient le premier signal et inversement). Ensuite, la différence entre ces deux multiplications est prise en compte et le résultat donne la vitesse du mouvement. La réponse du détecteur dépend de la phase, du contraste et de la vitesse du stimulus. De nombreux détecteurs réglés à différentes vitesses sont nécessaires pour coder la vitesse réelle du motif. La preuve expérimentale la plus convaincante de ce type de détecteur provient d'études sur la discrimination de la direction de cibles à peine visibles.
Filtrage d'énergie de mouvement
Le filtre d’énergie de mouvement est un modèle de capteurs de mouvement basé sur le principe des filtres invariants de phase. Ce modèle construit des filtres spatio-temporels orientés dans l'espace-temps pour correspondre à la structure des motifs en mouvement. Il est constitué de filtres séparables, pour lesquels les profils spatiaux conservent la même forme dans le temps mais sont mis à l'échelle par la valeur des filtres temporels. Les filtres d'énergie de mouvement correspondent à la structure des motifs en mouvement en ajoutant des filtres séparables. Pour chaque direction de mouvement, deux filtres espace-temps sont générés: un filtre symétrique et un filtre asymétrique. La somme des carrés de ces filtres est appelée énergie de mouvement. La différence de signal entre les deux directions s'appelle l'énergie adverse. Ce résultat est ensuite divisé par la sortie au carré d'un autre filtre, qui est réglé sur le contraste statique. Cette division est effectuée pour prendre en compte l'effet de contraste dans le mouvement. Les filtres d'énergie de mouvement peuvent modéliser un certain nombre de phénomènes de mouvement, mais ils produisent une mesure indépendante de la phase, qui augmente avec la vitesse mais ne donne pas une valeur fiable de la vitesse.
Gradients spatio-temporels

Ce modèle de capteurs de mouvement a été développé à l'origine dans le domaine de la vision par ordinateur. Il est basé sur le principe que le rapport de la dérivée temporelle de la luminosité de l'image sur la dérivée spatiale de la luminosité de l'image correspond à la vitesse de déplacement. Il est important de noter que dans les pics et les creux de l'image, ce modèle ne calculera pas une réponse adéquate, car la dérivée du dénominateur serait zéro. Afin de résoudre ce problème, les dérivées spatiales du premier ordre et d'ordres supérieurs en termes d'espace et de temps peuvent également être analysées. Les gradients spatio-temporels sont un bon modèle pour déterminer la vitesse de déplacement en tout point de l'image.
Les capteurs de mouvement sont sélectifs en orientation
[modifier | modifier le wikicode]L'une des propriétés des capteurs de mouvement est la sélectivité par rapport à l'orientation, qui limite l'analyse de mouvement à une seule dimension. Les capteurs de mouvement ne peuvent enregistrer le mouvement que dans une dimension le long d'un axe orthogonal à l'orientation préférée du capteur. Un stimulus contenant des caractéristiques d’une seule orientation ne peut se déplacer que dans une direction orthogonale à l’orientation du stimulus. Les signaux de mouvement unidimensionnels fournissent des informations ambiguës sur le mouvement des objets bidimensionnels. Une deuxième étape d'analyse du mouvement est nécessaire afin de déterminer la direction réelle du mouvement d'un objet ou modèle à deux dimensions. Les signaux de mouvement 1-D provenant de capteurs réglés selon différentes orientations sont combinés pour produire un signal de mouvement 2-D non ambigu. L'analyse du mouvement 2D dépend des signaux des capteurs locaux à orientation large ainsi que des signaux des capteurs à orientation étroite.
Suivi de charactéristiques
[modifier | modifier le wikicode]Une autre façon de percevoir le mouvement consiste à utiliser le suivi de charactéristiques. Le suivi de charactéristiques consiste à analyser si les caractéristiques locales d'un objet ont changé de position et à en déduire un mouvement. Dans cette section, certaines fonctionnalités des suivis de fonctionnalités sont mentionnées.
Les outils de suivi de charactéristiques échouent lorsqu'un mouvement-stimulus se produit très rapidement. Par rapport aux capteurs de mouvement, les détecteurs de suivi de charactéristiques ont l'avantage qu'ils peuvent percevoir le mouvement d'un objet même si le mouvement est interrompu par des intervalles intermittents sans réel signal. Ils peuvent également séparer ces deux phases (mouvements et intervalles vides). Les capteurs de mouvement, d'autre part, intégreraient simplement les phases vides avec le stimulus en mouvement et observeraient un mouvement continu. Les détecteurs pour suivi de charactéristiques opèrent sur la localisation des charactéristiques identifiées. Pour cette raison, ils ont un seuil de distance minimum qui correspond à la précision avec laquelle les emplacements des charactéristiques peuvent être distingués.
Les détecteurs pour suivi de charactéristiques ne montrent pas les effets secondaires de mouvement, qui sont des illusions visuelles causées par une adaptation visuelle. Les effets secondaires de mouvement se produisent lorsque, après avoir observé un stimulus en mouvement, un objet immobile semble se déplacer dans la direction opposée de celle du stimulus en mouvement observé précédemment. Il est impossible pour ce mécanisme de surveiller plusieurs mouvements simultanément et dans différentes parties du champ visuel. D'autre part, les mouvements multiples ne posent pas de problème aux capteurs de mouvement, car ils fonctionnent en parallèle sur tout le champ visuel.
Des expériences ont été menées en utilisant les informations susmentionnées pour parvenir à des conclusions intéressantes sur les détecteurs de suivi de caractéristiques. Des expériences avec de brefs stimuli ont montré que les motifs de couleur et les motifs de contraste très contrastés ne sont pas perçus par les suiveurs de caractéristiques, mais par les capteurs de mouvement. Des expériences avec des phases sans signal ont confirmé que le suivi de charactéristiques peut se produire avec des intervalles sans signal dans l'affichage. Ce n'est qu'à contraste élevé que les capteurs de mouvement perçoivent le mouvement de stimuli chromatiques et de stimuli de motifs de contraste. À faible contraste, les détecteurs analysent le mouvement des motifs chromatiques et des enveloppes de contraste et, à fort contraste, les capteurs de mouvement analysent les enveloppes de contraste. Les expériences dans lesquelles les sujets jugent plusieurs mouvements suggèrent que le suivi des caractéristiques est un processus qui se déroule sous contrôle conscient et que c'est le seul moyen d'analyser le mouvement des enveloppes de contraste dans des conditions à faible contraste. Ces résultats sont cohérents avec l’idée selon laquelle le mouvement des enveloppes et des motifs de couleur contrastés dépend du suivi des caractéristiques, sauf lorsque les couleurs sont bien supérieures au seuil de détection ou que le contraste moyen est élevé. La principale conclusion de ces expériences est que le suivi des caractéristiques permet probablement de percevoir les enveloppes de contraste et les motifs de couleur.
Illusions de mouvement
[modifier | modifier le wikicode]

En conséquence du processus de détection de mouvement, certaines images statiques peuvent nous sembler bouger. Ces images donnent un aperçu des hypothèses exprimées par le système visuel et sont appelées illusions visuelles.
Une illusion de mouvement célèbre liée aux signaux de mouvement de premier ordre est le phénomène Phi, qui est une illusion d'optique qui nous fait percevoir un mouvement au lieu d'une séquence d'images. Cette illusion de mouvement nous permet de regarder des films comme un continuum et non comme des images séparées. Le phénomène phi permet à un groupe d'images figées qui sont modifiées à une vitesse constante d'être considéré comme un mouvement constant. Le phénomène Phi ne doit pas être confondu avec le mouvement Bêta, car il s’agit d’un mouvement apparent provoqué par des impulsions lumineuses successives, tandis que le dernier est un mouvement apparent causé par des impulsions stationnaires lumineuses.
Les illusions de mouvement se produisent lorsque la perception du mouvement, l'analyse du mouvement et l'interprétation de ces signaux sont trompeuses et que notre système visuel crée des illusions concernant le mouvement. Ces illusions peuvent être classées selon le processus qui les cause. Les illusions sont classées comme des illusions liées à la détection de mouvement, à l'intégration 2D et à l'interprétation 3D.
Les illusions les plus courantes concernant la détection de mouvement sont les mouvements à quatre temps, les «Random Dot Kinetmatograms» et les illusions de signaux de mouvement de second ordre. Les illusions de mouvement les plus populaires en matière d'intégration 2D sont «Motion Capture», «Plaid Motion» et «Direct Repulsion». De la même manière, celles qui concernent l'interprétation 3D sont les suivantes: «Transformational Motion», «Kinetic Depth», «Shadow Motion», «Biological Motion», «Stereokinetic motion», «Implicit Figure Motion» et la «2 Stroke Motion». Il y a beaucoup plus d'illusions de mouvement, et elles montrent toutes quelque chose d'intéressant concernant les mécanismes de détection, de perception et d'analyse du mouvement humain. Pour plus d'informations, visitez le lien suivant: http://www.lifesci.sussex.ac.uk/home/George_Mather/Motion/
Problèmes ouverts
[modifier | modifier le wikicode]Bien que nous ne comprenions toujours pas la plupart des spécificités de la perception du mouvement, comprendre les mécanismes par lesquels le mouvement est perçu ainsi que les illusions de mouvement peuvent donner au lecteur un bon aperçu de l’état de l’art dans le sujet. Certains des problèmes ouverts concernant la perception du mouvement sont les mécanismes de formation des images 3D dans le mouvement global et le problème de l'ouverture.
Les signaux de mouvement global de la rétine sont intégrés pour aboutir à un signal de mouvement global à 2 dimensions; cependant, on ignore comment se forme le mouvement global 3D. Le problème de l'ouverture se produit parce que chaque champ réceptif dans le système visuel ne couvre qu'une petite partie du monde visuel, ce qui crée des ambiguïtés dans la perception. Le problème de l'ouverture concerne le problème qu'un contour en mouvement qui correspond à différentes possibilités de mouvement, lorsqu'il est observé localement. Cette ambiguïté est d'origine géométrique - les mouvements parallèles au contour ne peuvent pas être détectés, car les modifications apportées à cette composante du mouvement ne modifient pas les images observées par l'ouverture. La seule composante pouvant être mesurée est la vitesse orthogonale à l’orientation du contour; pour cette raison, la vitesse du mouvement pourrait être quelque chose de la famille des mouvements le long d'une ligne dans l'espace de la vitesse. Ce problème de l’ouverture n’est pas seulement observé dans les contours droits, mais également dans les contours aux courbes douces, puisqu’ils sont approximativement rectilignes lorsqu’ils sont observés localement. Bien que les mécanismes permettant de résoudre le problème d’ouverture soient encore inconnus, il existe certaines hypothèses sur la manière de le résoudre. Par exemple, il pourrait être possible de résoudre ce problème en combinant des informations sur l’espace ou à partir de contours différents du même objet.
Conclusions
[modifier | modifier le wikicode]Dans ce chapitre, nous avons présenté la perception du mouvement et les mécanismes par lesquels notre système visuel détecte les mouvements. Les illusions de mouvement ont montré à quel point les signaux de mouvement peuvent être trompeurs et conduire par conséquent à des conclusions erronées sur le mouvement. Il est important de se rappeler que la perception du mouvement et l'analyse du mouvement ne sont pas identiques. Les capteurs de mouvement et les détecteurs de suivi de charactéristiques se complètent pour que le système visuel perçoive le mouvement.
La perception du mouvement est complexe et reste un domaine de recherche ouvert. Ce chapitre décrit des modèles de fonctionnement des capteurs de mouvement et des hypothèses sur les caractéristiques des détecteurs de suivi de charactéristiques. Cependant, davantage d'expériences sont nécessaires pour connaître les caractéristiques de ces mécanismes et pour pouvoir construire des modèles plus proches des processus réels du système visuel.
La variété des mécanismes d'analyse et de perception du mouvement décrits dans ce chapitre, ainsi que la sophistication des modèles artificiels conçus pour les décrire, montrent qu'il existe une grande complexité dans la manière dont le cortex traite les signaux provenant de l'environnement extérieur. Des milliers de neurones spécialisés intègrent et interprètent des fragments de signaux locaux pour former des images globales d'objets en mouvement dans notre cerveau. Comprendre que tant d'acteurs et de processus dans notre corps doivent travailler de concert pour percevoir le mouvement rend notre capacité à le faire d'autant plus remarquable que nous humains sommes capables de le faire avec une telle facilité.
Perception des couleurs
[modifier | modifier le wikicode]Introduction
[modifier | modifier le wikicode]Les humains (ainsi que les primates comme les singes et les gorilles) ont la meilleure perception des couleurs chez les mammifères [1]. Ce n’est donc pas un hasard si la couleur joue un rôle important dans de nombreux domaines. Par exemple, la couleur est utile pour distinguer et différencier des objets, des surfaces, des paysages naturels et même des visages [2],[3]. La couleur est également un outil important pour la communication non verbale, y compris celle de l'émotion [4].
Pendant des décennies, il a été difficile de trouver le lien entre les propriétés physiques de la couleur et ses qualités perceptuelles. Celles-ci sont généralement étudiées sous deux approches différentes: la réponse comportementale causée par la couleur (également appelée réponse psychophysique) et la réponse physiologique réelle provoquée par celle-ci [5].
Ici, nous ne nous concentrerons que sur ce dernier aspect. L'étude des bases physiologiques de la vision des couleurs, sur laquelle on ne connaissait pratiquement rien avant la seconde moitié du XXe siècle, a progressé lentement et régulièrement depuis 1950. Des progrès importants ont depuis été réalisés dans de nombreux domaines, notamment au niveau des récepteurs. Grâce aux méthodes de biologie moléculaire, il a été possible de révéler des détails auparavant inconnus concernant la base génétique des pigments des cônes. De plus, de plus en plus de régions corticales sont influencées par des stimuli visuels, bien que la corrélation entre la perception des couleurs et l'activité physiologique dépendante de la longueur d'onde au-delà des récepteurs ne soit pas aussi facile à discerner [6].
Dans ce chapitre, notre objectif est d'expliquer les bases des différents processus de perception des couleurs le long du trajet visuel, de la rétine dans l'œil au cortex visuel dans le cerveau. Pour les détails anatomiques, veuillez vous référer à la section "Anatomie du système visuel" de ce Wikibook.
Perception de la couleur à la rétine
[modifier | modifier le wikicode]Toutes les couleurs pouvant être distinguées par les humains peuvent être produites par le mélange de seulement trois couleurs primaires (de base). Inspiré par cette idée de mélange des couleurs, il a été proposé de conserver la couleur au moyen de trois classes de capteurs, chacun ayant une sensibilité maximale à une partie différente du spectre visible [1]. Il a été explicitement proposé pour la première fois en 1853 qu’il existe trois degrés de liberté dans l’appariement normal des couleurs [7]. Ceci a été confirmé par la suite en 1886 [8] (avec des résultats remarquablement proches à ceux d'études récentes [9], [10]).
Ces capteurs de couleur proposés sont en réalité des cônes (remarque: dans ce chapitre, nous ne traiterons que les cônes. Les bâtonnets ne contribuent à la vision que lorsque la luminosité est faible. Bien qu'ils soient connus pour avoir un effet sur la perception des couleurs, leur influence est très faible et peut être ignorée ici)[11]. Les cônes font partie des deux types de cellules photoréceptrices présentes dans la rétine, avec une concentration significative dans la fovéa. Le tableau ci-dessous répertorie les trois types de cellules coniques. Ceux-ci se distinguent par différents types de pigments à la rhodopsine. Les courbes d'absorption correspondantes sont illustrées dans la figure ci-dessous.
Table 1: Aperçu général des types de cônes trouvés dans la rétine. Name Plus grande sensibilité à la couleur Courbe d'absorption maximale [nm]
| Nom | Plus grande sensibilité à la couleur | Courbe d'absorption maximale [nm] |
|---|---|---|
| S, SWS, B | Bleue | 420 |
| M, MWS, V | Verte | 530 |
| L, LWS, R | Rouge | 560 |
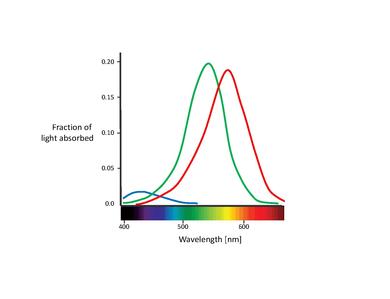
Bien qu'aucun consensus n'ait été atteint pour nommer les différents types de cônes, les désignations les plus utilisées font référence à leur pic de spectre d'action ou à la couleur à laquelle ils sont sensibles (rouge, verte, bleue)[6]. Dans ce texte, nous utiliserons la désignation S-M-L (pour longueur d'onde courte, moyenne et longue), car ces noms sont plus descriptifs. La nomenclature bleu-vert-rouge est quelque peu trompeuse, car tous les types de cônes sont sensibles à une large gamme de longueurs d'onde.
Une caractéristique importante des trois types de cônes est leur distribution relative dans la rétine. Il s'avère que les cônes S présentent une concentration relativement faible à travers la rétine, étant totalement absents dans la zone la plus centrale de la fovéa. En réalité, ils sont trop espacés pour jouer un rôle important dans la vision spatiale, bien qu'ils soient capables de médier une perception de frontière faible [12]. La fovéa est dominée par les cônes L et M. La proportion de ces deux derniers est généralement mesurée sous forme de ratio. Différentes valeurs ont été rapportées pour le rapport L / M, allant de 0,67 à 2 [13][14], ce dernier étant le plus accepté. La raison pour laquelle les cônes L sont presque toujours plus nombreux que les cônes M reste unconnu. Étonnamment, la ratio de cônes n’a presque aucun impact significatif sur la vision des couleurs. Ceci montre clairement que le cerveau est plastique, capable de donner un sens à tout signal de cône reçu [15], [16].
Il est également important de noter le chevauchement des spectres d'absorption des cônes L et M. Alors que le spectre d'absorption du cône S est clairement séparé, les pics des cônes L et M ne sont séparés que d'environ 30 nm, leurs courbes spectrales se chevauchant également de manière significative. Il en résulte une forte corrélation entre les captures de photons de ces deux classes de cônes. Cela s'explique par le fait que, pour obtenir la plus grande acuité possible au centre de la fovéa, le système visuel traite les cônes L et M de manière égale, sans tenir compte de leurs spectres d'absorption. Par conséquent, tout type de différence entraîne une détérioration du signal de luminance [17]. En d'autres termes, la faible séparation entre les spectres des cônes L et M pourrait être interprétée comme un compromis entre le besoin de vision couleur à contraste élevé et de vision en luminance à haute acuité. Ceci est congruent avec le manque de cônes S dans la partie centrale de la fovéa, où l'acuité visuelle est la plus élevée. En outre, l’espacement étroit des spectres d’absorption des cônes L et M pourrait également être expliqué par leur origine génétique. On suppose que les deux types de cônes ont évolué "récemment" (il y a environ 35 millions d'années) à partir d'un ancêtre commun, tandis que les cônes S se sont probablement séparés du récepteur ancestral beaucoup plus tôt [11].
Les fonctions d'absorption spectrale des trois types différents de cellules coniques sont la marque de la vision des couleurs humaine. Cette théorie a résolu un problème connu de longue date: bien que nous puissions voir des millions de couleurs différentes (les humains peuvent distinguer entre 7 et 10 millions de couleurs différentes [5]), nos rétines ne disposent tout simplement pas de suffisamment d'espace pour accueillir un détecteur individuel pour chaque couleur à chaque emplacement rétinien.
De la rétine au cerveau
[modifier | modifier le wikicode]Les signaux qui sont transmis de la rétine à des niveaux plus élevés ne sont pas de simples représentations ponctuelles des signaux des récepteurs, mais consistent plutôt en une combinaison sophistiquée de ces derniers. L'objectif de cette section est de fournir un résumé des chemins empruntés par certaines de ces informations.
Une fois que l'image optique sur la rétine est convertie en signaux chimiques et électriques dans les photorécepteurs, les signaux modulés en amplitude sont convertis en représentations modulées en fréquence aux niveaux de la cellule ganglionnaire et des niveaux supérieurs. Dans ces cellules neuronales, la magnitude du signal est représentée en termes de fréquence de pointes de tension déclenchées par la cellule, plutôt que par la différence de tension à travers la membrane cellulaire. Afin d'expliquer et de représenter les propriétés physiologiques de ces cellules, nous trouverons le concept de champs récepteurs très utile.
Un champ récepteur est une représentation graphique de la zone du champ visuel à laquelle une cellule donnée répond. De plus, la nature de la réponse est généralement indiquée pour diverses régions du champ récepteur. Par exemple, nous pouvons considérer le champ récepteur d'un photorécepteur comme une petite zone circulaire représentant la taille et l'emplacement de la sensibilité de ce récepteur particulier dans le champ visuel. La figure ci-dessous montre des exemples de champs récepteurs pour les cellules ganglionnaires, typiquement dans un antagonisme centre-pourtour. Le champ réceptif de gauche sur la figure illustre une réponse centrale positive (appelée centre ON). Ce type de réponse est généralement généré par une entrée positive d'un seul cône entouré d'une réponse négative générée par plusieurs cônes voisins. Par conséquent, la réponse de cette cellule ganglionnaire serait constituée des entrées de divers cônes présentant des signes à la fois positifs et négatifs. De cette manière, la cellule ne réagit pas seulement aux points de lumière, elle sert également de détecteur de bord (ou plus exactement de spot). Par analogie avec la terminologie de la vision par ordinateur, nous pouvons considérer les réponses des cellules ganglionnaires comme le résultat d’une convolution avec un noyau à détecteur de bord. Le champ de réception de droite de la figure illustre une réponse centrale négative (dite centre OFF), qui est tout aussi probable. Habituellement, les cellules centre ON et centre OFF apparaissent au même emplacement spatial, alimentées par les mêmes photorécepteurs, ce qui se traduit par une plage dynamique améliorée.
La figure inférieure montre qu’outre l’antagonisme spatial, les cellules ganglionnaires peuvent également avoir une opposition spectrale. Par exemple, la partie gauche de la figure du bas illustre une réponse rouge-verte adverse, le centre étant alimenté par une entrée positive provenant d'un cône L et l'entourage environnant étant alimenté par une entrée négative provenant de cônes M. En revanche, la partie droite de la figure du bas illustre la version centre OFF de cette cellule. Par conséquent, avant même que les informations visuelles ne quittent la rétine, un traitement a déjà eu lieu, avec un effet profond sur l'apparence de la couleur. Il existe d'autres types et variétés de réponses de cellules ganglionnaires, mais ils partagent tous ces concepts de base.



En route vers le cortex visuel primaire, les axones des cellules ganglionnaires se rassemblent pour former le nerf optique, qui fait saillie vers le corps géniculé latéral (CGL) du thalamus. Le codage dans le nerf optique est très efficace, réduisant au minimum le nombre de fibres nerveuses (limité par la taille du nerf optique) et gardant la taille de la tache aveugle de la rétine aussi petite que possible (environ 5 ° de large et 7 ° de haut). De plus, les cellules ganglionnaires présentées n'auraient aucune réponse à une illumination uniforme, car les zones positives et négatives sont équilibrées. En d'autres termes, les signaux transmis ne sont pas corrélés. Par exemple, les informations provenant de parties voisines de scènes naturelles sont fortement corrélées spatialement et donc très prévisibles [18]. L'inhibition latérale entre les cellules ganglionnaires rétiniennes voisines minimise cette corrélation spatiale, améliorant ainsi l'efficacité. Nous pouvons voir cela comme un processus de compression d'image réalisé dans la rétine.
Compte tenu du chevauchement des spectres d'absorption des cônes L et M, leurs signaux sont également fortement corrélés. Dans ce cas, l'efficacité du codage est améliorée en combinant les signaux de cône afin de minimiser ladite corrélation. Nous pouvons comprendre cela plus facilement en utilisant l'analyse en composantes principales (ACP). ACP est une méthode statistique utilisée pour réduire la dimensionnalité d'un ensemble de variables en transformant les variables d'origine en un ensemble de nouvelles variables, les composantes principales (CP). La première CP compte pour un montant maximal de la variance totale dans les variables d'origine, la deuxième CP compte pour un montant maximal de la variance non prise en compte par la première composante, etc. De plus, les CP sont linéairement indépendantes et orthogonales les unes aux autres dans l'espace des paramètres. Le principal avantage de l’ACP est que seules quelques-unes des CP les plus puissantes suffisent à couvrir la grande majorité de la variabilité du système [19]. Ce schéma a été utilisé avec les fonctions d'absorption du cône [20] et même avec les spectres naturels [21],[22]. Les CP détectées dans l'espace des excitations de cônes produites par des objets naturels sont 1) un axe de luminance où les signaux des cônes L et M sont ajoutés (L + M), 2) la différence des signaux des cônes L et M (L - M), et 3) un axe de couleur où la somme des signaux des cônes L et M est soustraite du signal du cône S (S - (L + M)). Ces canaux, issus d'une approche mathématique / informatique, coïncident avec les trois canaux rétino-géniculaires découverts lors d'expériences électrophysiologiques [23],[24]. En utilisant ces mécanismes, les informations visuelles redondantes sont éliminées dans la rétine.
Il existe trois canaux d’information qui communiquent effectivement cette information de la rétine à travers les cellules ganglionnaires au CGL. Ils diffèrent non seulement par leurs propriétés chromatiques, mais également par leur substrat anatomique. Ces canaux posent des limitations importantes pour les tâches concernant les couleurs de base, telles que la détection et la discrimination.
Dans le premier canal, la sortie des cônes L et M est transmise de manière synergique afin de diffuser les cellules bipolaires puis les cellules des couches magnocellulaires (M-) du CGL (à ne pas confondre avec les cônes M de la rétine)[24]. Les champs récepteurs des cellules M sont composés d'un centre et d'un pourtour, antagonistes dans l'espace. Les cellules M ont une sensibilité au contraste élevée pour les stimuli de luminance, mais elles ne montrent aucune réponse aux combinaisons des entrées adverses des récepteurs L-M [25].
Cependant, étant donné que les points nuls de différentes cellules M varient légèrement, la réponse de la population n’est jamais vraiment nulle. Cette propriété est en fait transmise aux zones corticales avec des entrées prédominantes de cellules M [26].
La voie parvocellulaire (P-) prend sa source dans les sorties individuelles des cellules bipolaires de type cône L ou M vers des «midget bipolar cells». Celles-ci fournissent des informations aux cellules P rétiniennes [11]. Dans la fovéa, les centres de champ récepteurs des cellules P sont formés par de simples cônes en L ou M. La structure du champ récepteur des cellules P est encore une question controversée. Cependant, la théorie la plus acceptée affirme que le pourtour est constitué d’un type de cône spécifique, ce qui crée un champ récepteur spatialement opposé aux stimuli de luminance [27]. Les couches parvocellulaires contribuent avec environ 80% du total des projections de la rétine au CGL [28].
Enfin, la voie koniocellulaire (K-) récemment découverte contient principalement des signaux provenant de cônes S [29]. Les groupes de ce type de cônes projettent dans des cellules bipolaires spéciales, qui à leur tour fournissent des informations à de petites cellules ganglionnaires spécifiques. Celles-ci ne sont généralement pas adverses dans l'espace. Les axones des petites cellules ganglionnaires projettent sur les couches minces du CGL (adjacentes aux couches parvocellulaires) [30].
Alors que les cellules ganglionnaires se terminent dans le CGL (en créant des synapses avec des cellules du CGL), il semble exister une correspondance biunivoque entre les cellules ganglionnaires et les cellules du CGL. Le CGL semble jouer le rôle de station relais pour les signaux. Cependant, il remplit probablement une fonction visuelle, car il existe des projections neuronales du cortex vers le CGL qui pourraient servir de mécanisme de rétroaction de commutation ou d’adaptation. Les axones des cellules CGL projettent dans l'aire visuelle 1 (V1) du cortex visuel dans le lobe occipital.
Perception de la couleur au cerveau
[modifier | modifier le wikicode]Dans le cortex, les projections des voies magno-, parvo- et koniocellulaire se terminent dans différentes couches du cortex visuel primaire. Les fibres magnocellulaires innervent principalement les couches 4Cα et 6. Les neurones parvocellulaires projettent principalement sur 4Cβ et les couches 4A et 6. Les neurones koniocellulaires se terminent dans les parts riches en cytochrome oxydase (CO) des couches 1, 2 et 3 [31].
Une fois dans le cortex visuel, le codage de l'information visuelle devient beaucoup plus complexe. De la même manière que les sorties de divers photorécepteurs sont combinées et comparées pour produire des réponses de cellules ganglionnaires, les sorties de diverses cellules de CGL sont comparées et combinées pour produire des réponses corticales. Au fur et à mesure que les signaux progressent dans la chaîne de traitement cortical, ce processus se répète avec un niveau de complexité qui augmente rapidement au point que les champs récepteurs commencent à perdre leurs sens distincts. Cependant, certaines fonctions et certains processus ont été identifiés et étudiés dans des régions spécifiques du cortex visuel.
Dans la région V1 (cortex strié), les neurones à double opposition - les neurones dont les champs récepteurs sont à la fois chromatiquement et spatialement opposés aux régions allumées / éteintes d'un seul champ récepteur - comparent les signaux de couleur dans l'espace visuel [32]. Ils constituent entre 5 et 10% des cellules de la région V1. Leur grande taille et leur faible pourcentage correspondent à la résolution spatiale médiocre de la vision des couleurs [1]. De plus, ils ne sont pas sensibles à la direction des stimuli en mouvement (contrairement à certains autres neurones de la région V1) et ne contribuent donc probablement pas à la perception du mouvement [33]. Cependant, étant donné leur structure de champ réceptif spécialisée, ces types de cellules constituent la base neurale des effets de contraste de couleurs, ainsi qu'un moyen efficace de coder la couleur elle-même [34],[35]. D'autres cellules V1 répondent à d'autres types de stimuli, tels que des bords orientés, diverses fréquences spatiales et temporelles, des localisations spatiales particulières et des combinaisons de ces caractéristiques, entre autres. De plus, nous pouvons trouver des cellules qui combinent linéairement les entrées des cellules du CGL ainsi que des cellules qui effectuent une combinaison non linéaire. Ces réponses sont nécessaires pour prendre en charge des fonctionnalités visuelles avancées, telles que la couleur elle-même.

Moins est connu sur les propriétés chromatiques de neurones individuels dans la région V2 que sur celles dans la région V1. À première vue, il semble qu'il n'y ait pas de différences majeures de codage de couleur dans V1 et V2 [36]. Une exception à cette règle est l'émergence d'une nouvelle classe de cellules susceptibales à des combinaisons complexes de couleurs [37]. Par conséquent, il a été suggéré que la région V2 soit impliquée dans l’élaboration de la teinte. Toutefois, cela reste très controversé et n’a pas été confirmé.
Suivant le concept modulaire mis au point après la découverte de la dominance oculaire fonctionnelle dans la région V1 et compte tenu de la ségrégation anatomique entre les voies P, M et K (décrites dans la section portant sur l'anatomie du système visuel), il a été suggéré qu'un système spécialisé consacré à l'analyse des informations de couleur devrait exister dans le cortex [38]. La région V4 est la région qui a historiquement attiré le plus d’attention en tant que possible "zone de couleur" du cerveau. Cela est dû à une étude influente selon laquelle la région V4 contenait 100% de cellules sélectives pour la teinte [39]. Cependant, cette affirmation a été contestée par plusieurs études ultérieures, certaines rapportant même que seulement 16% des neurones de la région V4 présenteraient un réglage de la teinte [40]. Actuellement, le concept le plus accepté est que la région V4 contribue non seulement à la couleur, mais également à la perception, à l’attention visuelle et à la stéréopsie. De plus, des études récentes se sont concentrées sur d'autres régions du cerveau essayant de trouver la "zone de couleur" du cerveau, telles que TEO [41] et PITd [42]. La relation de ces régions entre elles est toujours débattue. Pour réconcilier la discussion, certains utilisent le terme cortex temporal postérieur inférieur (TIP) pour désigner la région comprenant V4, TEO et PITd [1].
Si la réponse corticale dans les cellules des régions V1, V2 et V4 est déjà une tâche très compliquée, le niveau de complexité des réponses visuelles complexes dans un réseau d'environ 30 zones visuelles est énorme. La figure 4 montre une petite partie de la connectivité des différentes zones corticales (et non des cellules) identifiées [43].
À ce stade, il devient extrêmement difficile d'expliquer la fonction des cellules corticales simples en termes simples. En fait, la fonction d’une seule cellule n’a peut-être pas de sens puisque la représentation des différentes perceptions doit être répartie dans des collections de cellules à travers le cortex.
Mécanismes d'adaptation à la vision des couleurs
[modifier | modifier le wikicode]Bien que les chercheurs aient essayé d'expliquer le traitement des signaux de couleur dans le système visuel humain, il est important de comprendre que la perception de la couleur n'est pas un processus fixe. En réalité, divers mécanismes dynamiques permettent d'optimiser la réponse visuelle en fonction de l'environnement de visualisation. Les mécanismes d’adaptation à la noirceur, à la lumière et aux couleurs sont particulièrement importants pour la perception des couleurs.
Adaptation à l'obscurité
[modifier | modifier le wikicode]L'adaptation à l'obscurité fait référence au changement de sensibilité visuelle qui se produit lorsque le niveau d'éclairage est diminué. La réponse du système visuel à un éclairage réduit doit devenir plus sensible, ce qui augmente sa capacité à produire une réponse visuelle significative même lorsque les conditions d'éclairage sont sous-optimales [44].
La figure 5 montre la récupération de la sensibilité visuelle après passage d'un niveau d'éclairage extrêmement élevé à l'obscurité totale [43]. Tout d'abord, les cônes deviennent progressivement plus sensibles, jusqu'à ce que la courbe se stabilise au bout de quelques minutes. Après environ 10 minutes, la sensibilité visuelle est à peu près constante. À ce stade, le système de barres, avec un temps de récupération plus long, a récupéré suffisamment de sensibilité pour surperformer les cônes et ainsi récupérer le contrôle de la sensibilité globale. La sensibilité de la tige s’améliore progressivement aussi, jusqu’à ce qu’elle devienne asymptotique au bout de 30 minutes environ. En d'autres termes, les cônes sont responsables de la récupération de sensibilité pendant les 10 premières minutes. Ensuite, les bâtonnets surpassent les cônes et acquièrent une sensibilité complète au bout de 30 minutes environ.
Ceci n’est qu’un des nombreux mécanismes neuronaux d’adaptation aux conditions d’éclairage obscur. Parmi les autres mécanismes neuronaux, on compte le réflexe de pupille bien connu, l'appauvrissement et la régénération des photopigments, le contrôle du gain dans les cellules rétiniennes et d'autres mécanismes de niveau supérieur, et l'interprétation cognitive, entre autres.
Adaptation à la lumière =
[modifier | modifier le wikicode]L'adaptation à la lumière est essentiellement le processus inverse de l'adaptation à l'obscurité. En fait, les mécanismes physiologiques sous-jacents sont les mêmes pour les deux processus. Cependant, il est important de les considérer séparément car leurs propriétés visuelles sont différentes.

L'adaptation à la lumière se produit lorsque le niveau d'éclairage est augmenté. Par conséquent, le système visuel doit devenir moins sensible afin de produire des perceptions utiles, compte tenu du fait qu'il y a beaucoup plus de lumière visible disponible. Le système visuel a une plage dynamique de sortie limitée disponible pour les signaux qui produisent nos perceptions. Cependant, le monde réel a des niveaux d’éclairage d’au moins 10 ordres de grandeur supplémentaires. Heureusement, nous avons rarement besoin de visualiser l’ensemble des niveaux d’illumination en même temps.
À des niveaux de lumière élevés, l'adaptation est obtenue par le blanchiment des photopigments. Cela permet de capturer les photons dans les récepteurs et prévient la saturation de la réponse des cônes à des arrière-plans clairs. Les mécanismes d'adaptation à la lumière se produisent principalement dans la rétine [45]. En fait, les changements de gain sont en grande partie spécifiques à un cône et l'adaptation regroupe les signaux sur des zones ne dépassant pas le diamètre des cônes individuels [46],[47]. Cela indique une localisation de l'adaptation de la lumière qui peut avoir lieu aussi tôt que dans les récepteurs. Cependant, il semble y avoir plus d’un site d’échelle de sensibilité. Certains changements de gain sont extrêmement rapides, alors que d’autres prennent quelques secondes, voire quelques minutes, pour se stabiliser [48]. Habituellement, l’adaptation à la lumière prend environ 5 minutes (six fois plus rapidement que l’adaptation à l’obscurité). Cela pourrait indiquer l'influence des sites post-réceptifs.
La figure 6 montre des exemples d'adaptation à la lumière [43]. Si nous n’utilisions qu’une seule fonction de réponse pour cartographier la large plage d'intensités dans la sortie du système visuel, nous n'aurions alors qu'une très petite plage à notre disposition pour une scène donnée. Il est clair qu'avec une telle fonction de réponse, le contraste perçu d'une scène donnée serait limité. De plus, la sensibilité visuelle aux changements s’en verrait fortement dégradée en raison de problèmes du rapport signal sur bruit. Ce cas est indiqué par la ligne pointillée. D'autre part, les lignes continues représentent des familles de réponses visuelles. Ces courbes cartographient la plage d’éclairage utile d’une scène donnée dans toute la plage dynamique de la sortie visuelle, produisant ainsi la meilleure perception visuelle possible pour chaque situation. L'adaptation à la lumière peut être considérée comme le processus de glissement de la courbe de réponse visuelle le long de l'axe du niveau d'éclairage jusqu'à ce que le niveau optimal soit atteint pour les conditions de visualisation données.
Adaptation chromatique
[modifier | modifier le wikicode]Le concept général d’adaptation chromatique consiste en la variation de la hauteur des courbes de sensibilité spectrale à trois cônes. Cet ajustement est dû au fait que l'adaptation à la lumière se produit indépendamment dans chaque classe de cônes. Une formulation spécifique de cette hypothèse est connue sous le nom d'adaptation de von Kries. Cette hypothèse énonce que la réponse d'adaptation a lieu dans chacun des trois types de cônes séparément et équivaut à multiplier leurs sensibilités spectrales fixes par une constante de mise à l'échelle [49]. Si les poids de mise à l'échelle (également appelés coefficients de von Kries) sont inversement proportionnels à l'absorption de la lumière par chaque type de cône (c'est-à-dire qu'une absorption plus faible nécessitera un coefficient plus élevé), la mise à l'échelle de von Kries maintient une réponse moyenne constante dans chaque classe de cônes. Cela fournit un mécanisme simple mais puissant pour maintenir la couleur perçue des objets malgré les changements d'éclairage. Dans un certain nombre de conditions différentes, la mise à l'échelle de von Kries permet de bien prendre en compte les effets de l'adaptation à la lumière sur la sensibilité aux couleurs et sur l'apparence percue de celles-ci [50],[51].
Le moyen le plus simple de visualiser l'adaptation chromatique consiste à examiner un objet blanc sous différents types d'éclairage. Par exemple, examinons un morceau de papier sous une lumière du jour, fluorescente ou incandescente. La lumière du jour contient beaucoup plus d'énergie de longueurs d'ondes courtes que la lumière fluorescente, et l'éclairage incandescent contient beaucoup plus d'énergie de longueurs d'ondes longues que la lumière fluorescente. Cependant, malgré les différentes conditions d'éclairage, le papier conserve approximativement son aspect blanc sous les trois sources lumineuses. En effet, le système du cône S devient relativement moins sensible à la lumière du jour (afin de compenser l’énergie additionnelle des ondes courtes) et le système du cône L devient relativement moins sensible sous un éclairage incandescent (afin de compenser l’énergie additionnelle des ondes longues)[43].
Sources
[modifier | modifier le wikicode]- ↑ 1,0 1,1 1,2 et 1,3 Conway, Bevil R, « Color vision, cones, and color-coding in the cortex », dans The neuroscientist, vol. 15, 2009, p. 274-290
- ↑ Russell, Richard and Sinha, Pawan}, « Real-world face recognition: The importance of surface reflectance properties », dans Perception, vol. 36, no 9, 2007
- ↑ Gegenfurtner, Karl R and Rieger, Jochem, « Sensory and cognitive contributions of color to the recognition of natural scenes », dans Current Biology, vol. 10, no 13, 2000, p. 805-808
- ↑ Changizi, Mark A and Zhang, Qiong and Shimojo, Shinsuke, « Bare skin, blood and the evolution of primate colour vision », dans Biology letters, vol. 2, no 2, 2006, p. 217-221
- ↑ 5,0 et 5,1 Giordano Beretta, Understanding Color, Hewlett-Packard,
- ↑ 6,0 et 6,1 Boynton, Robert M, « Color vision », dans Annual review of psychology, vol. 39, no 1, 1988, p. 69-100
- ↑ Grassmann, Hermann, « Zur theorie der farbenmischung », dans Annalen der Physik, vol. 165, no 5, 1853, p. 69-84
- ↑ Konig, Arthur and Dieterici, Conrad, « Die Grundempfindungen und ihre intensitats-Vertheilung im Spectrum », dans Koniglich Preussischen Akademie der Wissenschaften, 1886
- ↑ Smith, Vivianne C and Pokorny, Joel, « Spectral sensitivity of the foveal cone photopigments between 400 and 500 nm », dans Vision research, vol. 15, no 2, 1975, p. 161-171
- ↑ Vos, JJ and Walraven, PL, « On the derivation of the foveal receptor primaries », dans Vision Research, vol. 11, no 8, 1971, p. 799-818
- ↑ 11,0 11,1 et 11,2 Gegenfurtner, Karl R and Kiper, Daniel C, « Color vision », dans Neuroscience, vol. 26, no 1, 2003, p. 181
- ↑ Kaiser, Peter K and Boynton, Robert M, « Role of the blue mechanism in wavelength discrimination », dans Vision research, vol. 125, no 4, 1985, p. 523-529
- ↑ Paulus, Walter and Kroger-Paulus, Angelika, « A new concept of retinal colour coding », dans Vision research, vol. 23, no 5, 1983, p. 529-540
- ↑ Nerger, Janice L and Cicerone, Carol M, « The ratio of L cones to M cones in the human parafoveal retina », dans Vision research, vol. 32, no 5, 1992, p. 879-888
- ↑ Neitz, Jay and Carroll, Joseph and Yamauchi, Yasuki and Neitz, Maureen and Williams, David R, « Color perception is mediated by a plastic neural mechanism that is adjustable in adults », dans Neuron, vol. 35, no 4, 2002, p. 783-792
- ↑ Jacobs, Gerald H and Williams, Gary A and Cahill, Hugh and Nathans, Jeremy, « Emergence of novel color vision in mice engineered to express a human cone photopigment », dans Science, vol. 315, no 5819, 2007, p. 1723-1725
- ↑ Osorio, D and Ruderman, DL and Cronin, TW, « Estimation of errors in luminance signals encoded by primate retina resulting from sampling of natural images with red and green cones », dans JOSA A, vol. 15, no 1, 1998, p. 16-22
- ↑ Kersten, Daniel, « Predictability and redundancy of natural images », dans JOSA A, vol. 4, no 112, 1987, p. 2395-2400
- ↑ I. T. Jolliffe, Principal Component Analysis, Springer,
- ↑ Buchsbaum, Gershon and Gottschalk, A, « Trichromacy, opponent colours coding and optimum colour information transmission in the retina », dans Proceedings of the Royal society of London. Series B. Biological sciences, vol. 220, no 1218, 1983, p. 89-113
- ↑ Zaidi, Qasim, « Decorrelation of L-and M-cone signals », dans JOSA A, vol. 14, no 12, 1997, p. 3430-3431
- ↑ Ruderman, Daniel L and Cronin, Thomas W and Chiao, Chuan-Chin, « Statistics of cone responses to natural images: Implications for visual coding », dans JOSA A, vol. 15, no 8, 1998, p. 2036-2045
- ↑ Lee, BB and Martin, PR and Valberg, A, « The physiological basis of heterochromatic flicker photometry demonstrated in the ganglion cells of the macaque retina », dans The Journal of Physiology, vol. 404, no 1, 1998, p. 323-347
- ↑ 24,0 et 24,1 Derrington, Andrew M and Krauskopf, John and Lennie, Peter, « Chromatic mechanisms in lateral geniculate nucleus of macaque », dans The Journal of Physiology, vol. 357, no 1, 1984, p. 241-265
- ↑ Shapley, Robert, « Visual sensitivity and parallel retinocortical channels », dans Annual review of psychology, vol. 41, no 1, 1990, p. 635--658
- ↑ Dobkins, Karen R and Thiele, Alex and Albright, Thomas D, « Comparison of red--green equiluminance points in humans and macaques: evidence for different L: M cone ratios between species », dans JOSA A, vol. 17, no 3, 2000, p. 545-556
- ↑ Martin, Paul R and Lee, Barry B and White, Andrew JR and Solomon, Samuel G and Ruttiger, Lukas, « Chromatic sensitivity of ganglion cells in the peripheral primate retina », dans Nature, vol. 410, no 6831, 2001, p. 933-936
- ↑ Perry, VH and Oehler, R and Cowey, A, « Retinal ganglion cells that project to the dorsal lateral geniculate nucleus in the macaque monkey », dans Neuroscience, vol. 12, no 4, 1984, p. 1101--1123
- ↑ Casagrande, VA, « A third parallel visual pathway to primate area V1 », dans Trends in neurosciences, vol. 17, no 7, 1994, p. 305-310
- ↑ Hendry, Stewart HC and Reid, R Clay, « The koniocellular pathway in primate vision », dans Annual review of neuroscience, vol. 23, no 1, 2000, p. 127-153
- ↑ Callaway, Edward M, « Local circuits in primary visual cortex of the macaque monkey », dans Annual review of neuroscience, vol. 21, no 1, 1998, p. 47-74
- ↑ Conway, Bevil R, « Spatial structure of cone inputs to color cells in alert macaque primary visual cortex (V-1) », dans The Journal of Neuroscience, vol. 21, no 8, 2001, p. 2768-2783
- ↑ Horwitz, Gregory D and Albright, Thomas D, « Paucity of chromatic linear motion detectors in macaque V1 », dans Journal of Vision, vol. 5, no 6, 2005
- ↑ Danilova, Marina V and Mollon, JD, « The comparison of spatially separated colours », dans Vision research, vol. 46, no 6, 2006, p. 823-836
- ↑ Wachtler, Thomas and Sejnowski, Terrence J and Albright, Thomas D, « Representation of color stimuli in awake macaque primary visual cortex », dans Neuron, vol. 37, no 4, 2003, p. 681-691
- ↑ Solomon, Samuel G and Lennie, Peter, « Chromatic gain controls in visual cortical neurons », dans The Journal of neuroscience, vol. 25, no 19, 2005, p. 4779-4792
- ↑ David H Hubel, Eye, brain, and vision, Scientific American Library/Scientific American Books,
- ↑ Livingstone, Margaret S and Hubel, David H, « Psychophysical evidence for separate channels for the perception of form, color, movement, and depth », dans The Journal of Neuroscience, vol. 7, no 11, 1987, p. 3416-3468
- ↑ Zeki, Semir M, « Colour coding in rhesus monkey prestriate cortex », dans Brain research, vol. 53, no 2, 1973, p. 422-427
- ↑ Conway, Bevil R and Tsao, Doris Y, « Color architecture in alert macaque cortex revealed by fMRI », dans Cerebral Cortex, vol. 16, no 11, 2006, p. 1604-1613
- ↑ Tootell, Roger BH and Nelissen, Koen and Vanduffel, Wim and Orban, Guy A, « Search for color 'center(s)'in macaque visual cortex », dans Cerebral Cortex, vol. 14, no 4, 2004, p. 353-363
- ↑ Conway, Bevil R and Moeller, Sebastian and Tsao, Doris Y, « Specialized color modules in macaque extrastriate cortex », dans 560--573, vol. 56, no 3, 2007, p. 560-573
- ↑ 43,0 43,1 43,2 et 43,3 Mark D Fairchild, Color appearance models, John Wiley & Sons,
- ↑ Webster, Michael A, « Human colour perception and its adaptation », dans Network: Computation in Neural Systems, vol. 7, no 4, 1996, p. 587 - 634
- ↑ Shapley, Robert and Enroth-Cugell, Christina, « Visual adaptation and retinal gain controls », dans Progress in retinal research, vol. 3, 1984, p. 263-346
- ↑ Chaparro, A and Stromeyer III, CF and Chen, G and Kronauer, RE, « Human cones appear to adapt at low light levels: Measurements on the red-green detection mechanism », dans Vision Research, vol. 35, no 22, 1995, p. 3103-3118
- ↑ Macleod, Donald IA and Williams, David R and Makous, Walter, « A visual nonlinearity fed by single cones », dans Vision research, vol. 32, no 2, 1992, p. 347-363
- ↑ Mary Hayhoe, Adaptation mechanisms in color and brightness, Springer,
- ↑ David L MacAdam, Sources of Color Science, MIT Press,
- ↑ Webster, Michael A and Mollon, JD, « Colour constancy influenced by contrast adaptation », dans Nature, vol. 373, no 6516, 1995, p. 694-698
- ↑ Brainard, David H and Wandell, Brian A, « Asymmetric color matching: how color appearance depends on the illuminant », dans JOSA A, vol. 9, no 9, 1992, p. 1443-1448